DOI: 10.31038/AFS.2026811
Abstract
The conchocelis, the filamentous sporophyte generation of Bangiales, proliferates through tip growth in which only the tips of branch initial cells elongate and subsequently divide perpendicularly to the apical–basal axis to form filamentous structures. As was found in terrestrial plants, the tip growth of conchocelis in the marine red alga Pyropia yezoensis is regulated by auxin, actin polymerization, and phosphatidylinositol signaling. Although extracellular Ca2+ influx is critical for tip growth in terrestrial plants, little is known about its involvement in the tip growth of algae. We therefore investigated whether extracellular Ca2+ influx is required for tip growth in P. yezoensis conchocelis. Treatment of isolated single-celled conchocelis with a Ca2+ chelator, ethylene glycol tetraacetic acid, inhibited both the formation of branch initials and the tip growth of branches in a dose-dependent manner. These findings indicate that extracellular Ca2+ influx is indeed involved in the tip growth of conchocelis, suggesting that the basic regulatory mechanisms governing tip growth may be conserved between marine red algae and terrestrial plants. Further confirmation of this possibility will require the characterization of the spatiotemporal patterns of Ca2+ oscillations and F-actin accumulation, as well as the subcellular localization of phosphoinositides and their catabolic enzymes in the P. yezoensis conchocelis.
Keywords
Branch initial, Ca2+, Conchocelis, Extracellular influx, Pyropia yezoensis, Red alga, Tip growth
Introduction
Patterns of anisotropic cell growth give rise to the body shapes of terrestrial plants and multicellular algae [1,2]. Diffuse growth, which results in the expansion of cell surfaces [3,4], produces planar or foliose shape, whereas tip growth [5-7], characterized by directional and highly localized expansion at the apex, produces a filamentous body architecture. Algal cells grow anisotropically through both diffuse and tip growth. We previously demonstrated that tip growth produces the filamentous structures of the conchocelis (sporophyte) and conchosporangium (conchosporophyte) of the red alga Pyropia yezoensis [8-12], although regulatory mechanisms of tip growth in P. yezoensis has not yet been fully elucidated. Moreover, how diffuse growth produces a foliose shape in the thallus (gametophyte) of P. yezoensis remains largely unknown. Therefore, characterizing the contrasting mechanisms that regulate patterns of diffuse and tip growth is necessary to understand the differences in growth–morphology relationships between the two life-cycle generations of Bangiales.
The mechanisms that regulate diffuse and tip growth systems have been studied extensively in terrestrial plants. Cortical microtubules (MTs), actin filament (F-actin), Rho GTPases, the histone variant H2A.Z, gibberellin, and brassinosteroids have all been implicated in the regulation of diffuse growth [1,3,13] likewise, MTs, F-actin, Rho GTPases, Ca2+ influx, reactive oxygen species, phosphatidylinositol signaling, auxin, and jasmonate have been shown to regulate tip growth [7,13-21]. Although research on the regulation of diffuse growth in macroalgae has shown little progress, recent studies have provided basic information about the mechanisms regulating tip growth in red and brown macroalgae [2,11,12,22]. Indeed, we demonstrated that auxin, phosphoinositide turnover, and actin polymerization are involved in the regulation of tip growth in P. yezoensis [11,12], consistent with findings in terrestrial plants [13,17,18,21]. Although extracellular Ca2+ influx is required for tip growth in terrestrial plants [17-19], little is known about its role in the tip growth of macroalgae. We therefore sought to determine whether extracellular Ca2+ influx is required for tip growth in P. yezoensis conchocelis.
Materials and Methods
Single-celled conchocelis of P. yezoensis strain U-51 were used for these experiments, enabling us to visualize and quantitatively analyze the initiation and progression of tip growth in detail [11]. Conchocelis were maintained in artificial seawater at 15°C under 60 µE/m²/s of light with a short-day cycle (10-h light/14-h dark) [23]. Single-celled conchocelis were prepared as described in [11]. In brief, aggregates of multicellular conchocelis were chopped with a razor blade, and small fragments of conchocelis were separated from larger pieces by filtration through a 10-μm nylon mesh. The separated conchocelis were then incubated in 30 mL of artificial seawater [23] at 15°C for 10 min. Unbranched single-celled conchocelis (Figure 1) were identified by observation under an Olympus IX73 light microscope equipped with an Olympus DP22 camera (Olympus Corporation, Tokyo, Japan), drawn into a micropipette, and transferred to 96-well plates (one cell per well containing 200 μL of artificial seawater). Ethylene glycol tetraacetic acid (EGTA; Dojindo Laboratories, Japan), an effective Ca2+ chelator, was dissolved in artificial seawater to create a 0.5 M stock solution (adjusted to pH 8.0 with NaOH) and stored at −30°C before use. Single-celled conchocelis were treated with 0, 500, or 750 μM EGTA for 3 days, and the growth and morphology of the side branches were observed using the microscope described above.
Figure 1: Representative image of an isolated single-celled conchocelis. Nonbranched single-celled conchocelis were used for EGTA treatment experiments. Scale bar, 25 µm.
Results and Discussion
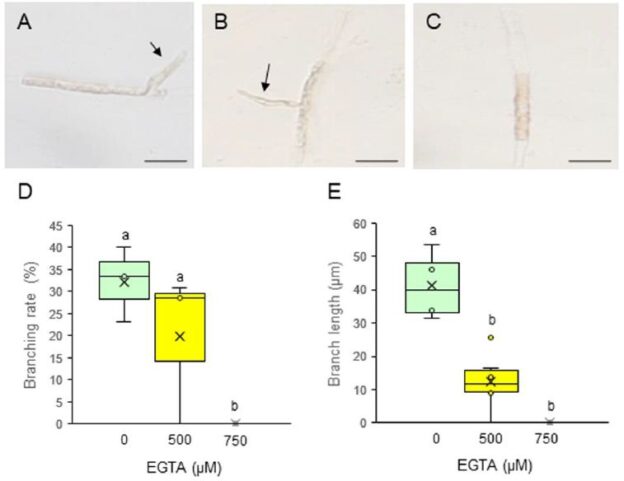
To evaluate the effects of EGTA on the initiation and elongation of side branches, we measured the branching rate (the percentage of observed cells that produced side branches) and branch length, respectively. Side branches were initiated and underwent elongation when treated with 0 (control) or 500 μM EGTA (Figure 2A and 2B), but 750 μM EGTA completely prevented both the initiation and growth of side branches (Figure 2C). The branching rate and branch length were affected by the presence of EGTA in a dose-dependent manner, being strongly inhibited at 750 μM (Figure 2D and 2E). These results demonstrate that extracellular Ca2+ influx is important for the production and tip growth of branch initials from differentiated conchocelis cells. It reminds to be elucidated whether washing out of EGTA and subsequent addition of CaCl2 in the medium recover branch initiation and branch growth and whether inhibition of channel-mediated Ca2+ influx by treatment with LaCl3 prevents tip growth.
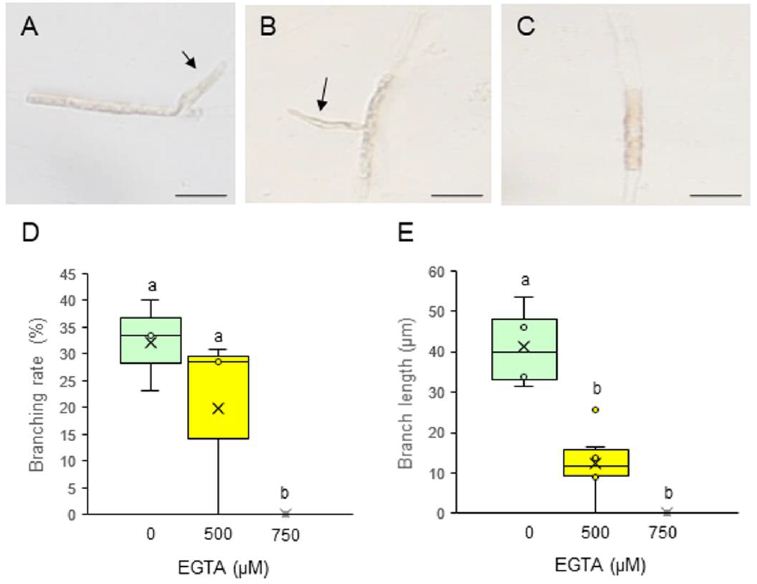
Figure 2: Effects of EGTA treatment on the tip growth of side branches from single-celled conchocelis. (A–C) Photographs of single-celled conchocelis treated with 0 (A), 500 (B), or 750 μM EGTA (C) for 3 days, for which each treatment was employed total 16 isolated cells for observation. Arrows indicate tip-growing branches initiated from single-celled conchocelis. Scale bars, 25 μm. (D, E) Branching rate (D) and branch length (E) after treatment of single-celled conchocelis with 0, 500, or 750 μM EGTA for 3 days. Center line, median; cross, mean; box limits, interquartile range with upper and lower quartiles; points, individual data points; whiskers, range with maximum and minimum values. Lowercase letters in (D) and (E) denote significant differences between treatments based on three independent experiments (n = 3) as determined by the Tukey–Kramer test (p < 0.05).
There are two important steps in the tip growth of conchocelis: the formation of branch initials in non-dividing differentiated cells (initiation of tip growth) and the polar directional growth of these initials to form elongated branches [12]. Because the treatment of single-celled conchocelis with EGTA impaired both branch initiation and branch growth, we concluded that Ca2+ is required for both critical steps in tip growth. To confirm this proposal, measurements of the time-course of both production of branch initial and its growth in single-celled conchocelis are necessary for elucidation of a question which step requires extracellular Ca2+ influx. In addition, we previously demonstrated that these two steps are regulated by auxin, phosphoinositide turnover, and actin polymerization [11,12]. These findings are consistent with our understanding of tip-growth regulation in the pollen tubes and root hairs of terrestrial plants [13,14,16-18,21]. It is therefore possible that the basic regulatory mechanisms governing tip growth are conserved between aquatic red algae and terrestrial plants. Factors such as Ca2+, enzymes related to phosphoinositide turnover, and F-actin act at the cell apex to enable the polar growth of filamentous tissues such as root hairs and pollen tubes [14,16,18]; however, little is known about the spatial distribution of these factors during the initiation and elongation of branches in P. yezoensis conchocelis. Therefore, determining the specific regions where auxin and Ca2+ exert their effects and characterizing the subcellular distributions of F-actin and enzymes involved in phosphoinositide turnover will be critical for fully understanding tip-growth regulation in conchocelis branches. A live-imaging technique for conchocelis tip growth has recently been reported [10]. Improvements to this system are expected to enable the visualization of Ca2+ influx, F-actin distribution, and the localization of phosphoinositides and related enzymes, providing spatiotemporal details of polarity establishment and tip growth in P. yezoensis conchocelis.
Despite progress in understanding tip growth in the conchocelis, much less is known about the regulation of diffuse growth that gives rise to the foliose thallus of the gametophyte. It is remarkable that foliose and filamentous generations survive independently in P. yezoensis and other Bangiales [9], suggesting differences in the regulatory mechanisms governing these two growth patterns. However, the shared involvement of MTs, F-actin, and Rho GTPases in both diffuse and tip growth of terrestrial plants [1] raises the question of how common factors can regulate different types of growth. Characterizing the contrasting regulatory mechanisms that govern diffuse and tip growth will be essential for understanding the physiological and molecular bases of the distinct multicellular body plans of heteromorphic generations in P. yezoensis.
Author Contributions
RI: Methodology, Investigation, Data curation, Formal analysis, Validation, Visualization.
KM: Conceptualization, Methodology, Formal analysis, Validation, Visualization, Supervision, Writing—original draft preparation and reviewing and editing, Funding acquisition.
Conflict of Interest
The authors declare no conflicts of interest. No aspects of the study required informed consent, and the work did not involve human or animal subjects. All authors have read and agreed to authorship and to the submission of the manuscript for peer review.
Data Availability Statement
Data are contained within the article.
Funding
This research was supported in part by the designated research fund from Miyagi University.
Acknowledgments
We are grateful to the Marine Resources Research Center of Aichi Fisheries Research Institute for kindly providing P. yezoensis strain U51.
References
- Guerriero G, Hausman JF, Cai G (2014) No stress! Relax! Mechanisms governing growth and shape in plant Int J Mol Sci 15(3): 5094-114. [crossref]
- Rabillé H, Billoud B, Tesson B, Le Panse S, Rolland É, et (2019) The brown algal mode of tip growth: Keeping stress under control. PLoS Biol 17(1): e2005258. [crossref]
- Braidwood L, Breuer C, Sugimoto K (2014) My body is a cage: mechanisms and modulation of plant cell New Phytol 201(2): 388-402. [crossref]
- Cosgrove DJ (2018) Diffuse growth of plant cell walls. Plant Physiol 176(1): 16-27. [crossref]
- Dumais J, Shaw SL, Steele CR, Long SR, Ray PM (2006) An anisotropic-viscoplastic model of plant cell morphogenesis by tip growth. Int J Dev Biol 50(2-3): 209-222.
- Rounds CM, Bezanilla M (2013) Growth mechanisms in tip-growing plant Annu Rev Plant Biol 64: 243-265. [crossref]
- Bibeau JP, Galotto G, Wu M, Tüzel E, Vidali L (2021) Quantitative cell biology of tip growth in Plant Mol Biol 107(4-5): 227-244. [crossref]
- Takahashi M, Mikami K (2016) Phototropism in the marine red macroalga Pyropia yezoensis. Am J Plant Sci 7: 2412-2428. https://doi.org/10.4236/ajps.2016.717211
- Mikami K, Li C, Irie Hama Y (2019) A unique life cycle transition in the red seaweed Pyropia yezoensis depends on apospory. Commun Biol 2: 299. [crossref]
- Hiwatashi Y, Shimada M, Mikami K, Takada N (2022) Establishment of a live-imaging analysis for polarized growth of conchocelis in the multicellular red alga Neopyropia yezoensis Front Plant Sci 12:716011. [crossref]
- Taya K, Takeuchi S, Takahashi M, Hayashi K, Mikami K (2022) Auxin regulates apical stem cell regeneration and tip growth in the marine red alga Neopyropia yezoensis. Cells 11(17): 2652. [crossref]
- Irie R, Mikami K (2025) Phosphoinositide signaling and actin polymerization are critical for tip growth in the marine red alga Pyropia yezoensis. Plants 14(14): 2194. [crossref]
- Ma C, Chang J, Hazelwood OS, Arif Ashraf M, Nan Q (2025) Establishing cell polarity in plants: the role of cytoskeletal structures and regulatory pathways. Front Cell Dev Biol 13: 1602463. [crossref]
- Ischebeck T, Seiler S, Heilmann I (2010) At the poles across kingdoms: phosphoinositides and polar tip Protoplasma; 240(1-4): 13-31. [crossref]
- Saavedra L, Mikami K, Malhó R, Sommarin M (2012) PIP kinases and their role in plant tip growing Plant Signal Behav 7(10): 1302-1305. [crossref]
- Qu X, Jiang Y, Chang M, Liu X, Zhang R, Huang S (2015) Organization and regulation of the actin cytoskeleton in the pollen Front Plant Sci 5: 786. [crossref]
- Lee YJ, Yang Z (2008) Tip growth: signaling in the apical Curr Opin Plant Biol 11(6): 662-671. [crossref]
- Scholz P, Anstatt J, Krawczyk HE, Ischebeck T (2020) Signalling pinpointed to the tip: the complex regulatory network that allows pollen tube growth. Plants 9: 1098. [crossref]
- Zhang X, Bian A, Li T, Ren L, Li L, Su Y, Zhang Q (2022) ROS and calcium oscillations are required for polarized root hair growth. Plant Signal Behav 17(1): 2106410. [crossref]
- Han X, Kui M, He K, Yang M, Du J, et (2023) Jasmonate-regulated root growth inhibition and root hair elongation. J Exp Bot 74: 1176-1185. [crossref]
- Berdion Gabarain V, Núñez-Lillo G, Pěnčík A, Carignani Sardoy M, Ibeas MA, et (2025) Enhanced auxin signaling promotes root-hair growth at moderately low temperature in Arabidopsis thaliana. Plant Commun 6(6): 101350. [crossref]
- Charrier B, Le Bail A, de Reviers B (2012) Plant Proteus: brown algal morphological plasticity and underlying developmental Trends Plant Sci 17(8): 468-77. [crossref]
- Li C, Ariga I, Mikami K (2019) Difference in nitrogen starvation-inducible expression patterns among phylogenetically diverse ammonium transporter genes in the red seaweed Pyropia yezoensis. Am J Plant Sci, 10: 1325–1349. https://doi: 4236/ajps.2019.108096