Abstract
Developing a storehouse of semantic or world knowledge is an important developmental achievement. In addition to direct learning such as through books and lectures, semantic knowledge accumulates through so-called productive processes, which permit going beyond the given to generate new knowledge. In the present research, we tested whether the specific productive process of self-derivation through memory integration makes unique contributions to semantic knowledge, even when other known or hypothesized correlates of knowledge (i.e., verbal and visual abilities) are considered. One-hundred-forty- eight 8-12-year-old children participated in two waves of testing, one year apart. At Wave 1, latent constructs of self-derivation and fact recall and verbal and visual abilities each predicted unique and overlapping variance in the latent construct, knowledge; total variance in knowledge accounted for R2 = .56. At Wave 2, verbal and visual abilities predicted unique variance in knowledge, and self-derivation and fact recall contributed to additional shared variance; total variance in knowledge accounted for R2 = .70. Longitudinally, both self-derivation and fact recall and verbal and visual abilities from Wave 1 predicted unique and overlapping variance in the latent construct, knowledge at Wave 2; total variance in knowledge accounted for R2 = .63. The work establishes self-derivation through memory integration as a mechanism of semantic knowledge development.
Keywords
development, longitudinal, productive processes, self-derivation, semantic knowledge
Developing a storehouse of knowledge about the world—so-called semantic knowledge—is a critical developmental achievement. As such, it is important to establish the correlates of the development of semantic knowledge. One contributor to semantic content is memory for information that is directly taught or learned, such as provided through lectures, books, videos, and museum exhibits. Other contributors include productive processes such as analogy, transitive and associative inference, and induction, to name a few. Productive processes go beyond what is given through direct experience, and therefore have the potential to accelerate accumulation of knowledge [1-6]. From among the larger suite of productive processes, self- derivation through memory integration has emerged as a valid model of knowledge accumulation[7,8]. Self-derivation through memory integration involves combination of separate yet related episodes of new learning and use of the novel combination as a basis for inference of new knowledge. For example, a learner may be taught that the heart is the only muscle that never tires, and that the only muscle that never tires is powered by electricity. Combination of these facts supports derivation of the new knowledge that the heart is powered by electricity. This new information can be incorporated into semantic knowledge, even though it was not directly experienced [9].
Consistent with the suggestion that self-derivation through memory integration is a valid model of knowledge accumulation, the process is both a concurrent and longitudinal predictor of academic achievement among elementary-school children, even when other known correlates of academic achievement (e.g., working memory) are considered [10,11]. It also is a concurrent and longitudinal predictor of knowledge in a number of domains as tested by the Woodcock- Johnson measures of achievement[12,13]. However, beyond the classroom, self-derivation through integration has not been tested against other known correlates of domain knowledge, such as existing knowledge as measured by vocabulary, and working memory. As a result, it has not been established as a unique predictor of knowledge accumulation. Accordingly, in the present research, we tested self- derivation through memory integration along with measures of other cognitive abilities, as concurrent and longitudinal predictors of semantic knowledge. The major goal was to determine whether self- derivation makes unique contributions to prediction and thus can be considered a mechanism of semantic knowledge development.
Self-derivation through Memory Integration
Self-derivation through memory integration involves combination of separate yet related facts that have been directly taught, to support derivation of new knowledge. With some important exceptions, the task is structured like those that test associative inference [3]. In associative inference tasks, participants may learn arbitrarily-related pairs of images or symbols that have an overlapping element. For example, they learn the image pairs zucchini-bucket (i.e., notation: A [for zucchini], B [for bucket]) and bucket-truck (B, C [for truck]). They typically are informed that they can form an indirect relation between the two image pairs, based on the overlapping element (bucket). They then are tested for the indirect relation between zucchini-truck (AC; [14]). As suggested by the opening example (above), the test of self- derivation through memory integration is structured similarly to tests of associative inference, with the important exceptions that (a) the stimuli are true facts expected to be unfamiliar to participants, (b) participants are not made aware that the facts they are learning are related to one another, and (c) nor are they made aware that they can form an indirect relation based on an overlapping element. In these respects, the self-derivation through integration task mimics learning outside the laboratory.
Self-derivation through memory integration occurs across ages, venues, and domains. In children as young as 4 years, new information is taught in the context of brief narratives, in the laboratory [15,16]; in kindergarten and elementary school classrooms [17]; or in children’s homes, in the course of shared book reading [18] or visits to virtual museum exhibits[19]. Among children 8 years and older and adults, new information is taught in individual sentences (such as the opening example, above). Information is drawn from a range of domains, including arts and humanities, math, science, and elementary [7] and college [20] curricula. Developmental differences in self-derivation through integration are readily apparent in early childhood. By roughly 8 years of age, developmental differences are eclipsed by individual differences that persist at least into young adulthood [21].
Self-derivation through Memory Integration and Academic Achievement
As expected from a model of learning, performance on tasks of self-derivation through memory integration relates to measures of academic achievement. For example, among children in Grades 1-3 (roughly ages 7-10 years), self-derivation through integration was found to predict both math and reading achievement, as measured by end-of-year assessments. It explained up to 23% of the variance in math and 12% in reading [17]. Similar findings were reported in [10], among children in 2nd and 3rd grades. Self-derivation through memory integration also predicts math and reading achievement over one year, among children in 2nd and 3rd grade at Time 1 and 3rd and 4th grade at Time 2 ([11], Study 2). Self-derivation through memory integration performance at Time 1 predicted 18% and 21% of the variance in the Time 2 end-of-year math and end-of-year reading achievement, respectively.
Of course, self-derivation through memory integration is not the only predictor of academic achievement. For example, a consistent correlate of academic achievement is working memory [22, 23]. Children’s working memory skills predict math and reading achievement 6 years later, above measures of IQ [24]. Working memory in preschoolers predicts math and reading achievement in the first three years of primary school, and visual short term memory predicts math achievement in particular[25]. Further, visuospatial working memory predicts student’s math achievement from first grade to second grade [26].
Importantly, self-derivation through memory integration is predictive of academic performance even when other known correlates of achievement are considered. Specifically,[10] tested the concurrent predictive utility of self-derivation through integration when considered along with verbal comprehension, nonverbal intelligence, and working memory and other executive functions. End-of-year math performance was jointly predicted by self-derivation through integration, verbal comprehension, nonverbal intelligence, and working memory. The total variance in math achievement accounted for was 51% and 41% among children in 2nd and 3rd grades, respectively. Self-derivation through integration alone accounted for 31% and 25% of the total for children in 2nd and 3rd grades, respectively. In the models predicting end-of-year reading performance, for children in 2nd grade, self-derivation through integration was the only significant predictor, accounting for 32% of the variance in reading achievement. For children in the 3rd grade, self-derivation through integration was a significant predictor, along with verbal comprehension and nonverbal intelligence. The full mode accounted for 48% of the variance in reading achievement; self-derivation through integration alone accounted for 22% of the total variance. Longitudinally, as noted above, Esposito & Bauer [11], Study 2) found that self-derivation through memory integration performance at Time 1 predicted 18% of the variance in the Time 2 end-of-year math achievement. The addition of measures of vocabulary and parent education in the model did not result in significant improvement. Thus self-derivation through integration was the sole longitudinal predictor of math achievement. For the Time 2 end-of-year assessment of reading achievement, self- derivation through memory integration at Time 1 predicted 21% of the variance. The addition to the model of vocabulary and parent education increased the variance accounted for to 31%.
Self-derivation through Memory Integration and Semantic Knowledge
Just as self-derivation through integration is not the only predictor of academic achievement, so too is it the case that academic achievement is not the only measure of semantic knowledge. Indeed, end-of-year achievement tests are administered en-mass in sometimes noisy and crowded classrooms and thus may not reflect a given student’s actual achievement. As well, they do not provide information about the broader contents of semantic knowledge, beyond the skills of math and reading.
As expected from a model of learning more broadly, performance on tasks of self-derivation through memory integration relates to individually administered tests of knowledge in a larger number and variety of domains, thus providing converging evidence of relations between self-derivation through integration and semantic knowledge. Specifically, Bauer and colleagues [12] tested the predictive utility of self-derivation through integration to six domains assessed by the Woodcock-Johnson IV tests of achievement [27]. The sample was 8- to 12-year-old children tested individually, on a Zoom platform. The specific domains of achievement tested were humanities, math, sciences, and social studies, as well as general knowledge of what and where, and passage comprehension [12]. The battery thus provided a relatively comprehensive assessment of the status of semantic knowledge.
In tests of concurrent relations, self-derivation through integration predicted 15%-36% of the variance in 8- to 12-year-old children’s performance across the domains [12]. The exception was the domain of sciences, for which self-derivation through integration was not a significant predictor. [13] extended the sample to examine longitudinal relations between self-derivation through integration and the knowledge domains in the same sample of children, across one year’s time. They combined the six individual domains into one latent factor, knowledge. Self-derivation through integration at Time 1 was a significant predictor of Time 2 knowledge. Self-derivation through memory integration at Time 1 was a stronger predictor of Time 2 knowledge (β = .43) than either age (β = .33) or memory for directly taught facts (β = .30). To date, the predictive utility of the productive process of self-derivation through integration has not been tested in the context of a larger suite of cognitive abilities known to be or logical predictors of learning and semantic knowledge. This test is the subject of the present research.
The Present Research
The major goal of the present research was to determine whether self-derivation makes unique contributions to prediction of domain knowledge and thus can be considered a mechanism of semantic knowledge development in childhood. We addressed the question concurrently and longitudinally over one year, in a sample of 148 8- to 12-year-old children. The sample was the same as tested by Bauer and colleagues [12,13]. In the prior studies, self-derivation was found to predict semantic knowledge both concurrently [12] and longitudinally [13]. In those studies, self-derivation through integration—and its attendant measures of memory for the directly taught facts on which self-derivation depends—were the only measures in the predictive models. The extension provided by the present research was to include in predictive models measures of a number of cognitive abilities that have been found to relate to self-derivation itself, both concurrently and longitudinally [8].
The specific cognitive abilities measured in the present research were (a) verbal comprehension (vocabulary, synonyms, antonyms, analogies), which often is considered a measure of semantic knowledge and even as a proxy for intelligence [28]; (b) working memory, a known correlate of academic achievement in childhood [22]; (c) visual-auditory learning, which is a measure of encoding and retention of new information, such as on which self-derivation through integration depends; and (d) visualization, a potential predictor of knowledge of math and sciences in particular. The measure of verbal comprehension was from the WJ-III; the measures of working memory, visual-auditory learning, and visualization were from the WJ-IV. In [12], verbal comprehension, visual-auditory learning, and visualization were significantly positively correlated with self-derivation through integration; working memory was not. However, they were neither concurrently nor longitudinally predictive of self-derivation when measures of directly taught facts—such as on which self-derivation through integration depends—were included in the model.
Consideration of self-derivation through integration and measures of other cognitive abilities as predictors of semantic knowledge across a number and variety of domains permits test of the major question motivating the research, namely, whether self-derivation is a unique predictor, concurrently and/or longitudinally. Based on findings from elementary classroom-based studies [10, 11], we expected self-derivation through integration to make unique contributions to variance in semantic knowledge. To our knowledge, beyond the tests to evaluate the psychometric properties of the Woodcock-Johnson measures, there have not been assessments of relations between the measures of cognitive abilities and those of semantic knowledge. The present research thus presents a unique test of this question, as well as of the question of unique prediction of the measures. The work stands as an evaluation of the potential of self-derivation through integration as a mechanism of semantic knowledge development.
Method
Participants
Participants were 148 children ages 8 to 12 years (M=10.45, SD=1.37, range=8.13-12.91 years) at enrollment. Nominally, there were 30 8-year-olds, 27 9-year-olds, 42 10-year-olds, 20 11-year-olds, and 29 12-year-olds. Most participants were recruited from a database of families who had expressed interest in taking part in research in child development. The rest were recruited through a marketing firm or by referral by participants already recruited (i.e., snowballing). The sample was 76 female (51%) and 70 male (47%); 2 caregivers did not report their child’s assigned sex at birth. The racial and ethnic composition of the sample was 7% Asian, 15% Black/African American, 1% Middle Eastern or Arab, 65% White or Caucasian, 9% multi-racial, and 3% did not report on their children’s race; 7% of the sample self-identified as Hispanic or Latinx. Ninety-two percent of caregivers had received at least some college education, and 56% of caregivers had received at least some graduate level education; 7% did not report caregiver education. All demographic information is based on caregiver report provided at Year 1 (analyses of demographic information are provided [12], and are not repeated in the present report). An additional 25 children were recruited but their data were not included because of failure to complete all four sessions across the two years of data collection (18), technical failure (3), prior participation in a related study (1), parental report of a developmental disability (2), and child-initiated request to end the session before all tasks were administered (1).
The study involved two sessions in each of two consecutive years. Within each year, the two sessions took place an average of 7 days apart (Year 1 range=6-13 days; Year 2 range=5-14 days). Children participated in Year 2 Session 1 an average of 364 days after completing Year 1 Session 1 (range=341–435 days). This study was begun during the 2020 COVID-19 related shutdown, and all data were collected online via Zoom. Written informed consent for children’s participation was provided by the children’s caregivers; children provided verbal assent. Participants were compensated with $40.00 in an e-gift card at the end of the second session at Year 1, and with $50.00 in an e-gift card at the end of the final session at Year 2. The procedures were reviewed and approved by the university Institutional Review Board.
Stimuli and Materials
At both waves of data collection, the full protocol included tests of self-derivation through memory integration, recall of directly-taught and of self-derived facts, measures of candidate component cognitive abilities, and measures of domain knowledge. Concurrent and longitudinal relations between self-derivation through integration and measures of domain knowledge were reported [12,13], respectively. The unique contribution of the present report is to augment measures from the self-derivation through integration task with measures of candidate component cognitive abilities as concurrent and longitudinal predictors of measures of domain knowledge. The current analyses thus permit examination of the unique and combined predictive utility of the entire suite of potential predictors of domain knowledge. The exception is the measure of recall of self-derived facts. As reported in [12], it was not predictive of domain knowledge and for this reason, it is not included in the present report.
Outcome Measures: Tests of Achievement/Semantic Knowledge
The outcome variable of interest was domain knowledge/ semantic knowledge. Six tests from the Woodcock-Johnson® IV (WJ® IV) Tests of Achievement and Tests of Cognitive Abilities [27] were used to measure semantic knowledge. The tests assess content knowledge in a number of different domains: (a) Test 2, Applied problems (analyze and solve math problems); (b) Test 4, Passage comprehension (use syntactic and semantic cues to provide a missing word in a text passage); (c) Tests 8a and 8b, General knowledge-what and General knowledge-where (respond to questions of the form “What would you do with a ?” and “Where would you find a ?”); (d) Test 18, Sciences (knowledge of anatomy, biology, geology, medicine, chemistry, and physics); (e) Test 19, Social Studies (knowledge of economics, psychology, government, history, and geography); and (f) Test 20, Humanities (knowledge of music, art, and literature). For children in the age range of 5 to 19 years, the tests have median reliability of .76 to .92. To accommodate online data collection, the tests were rendered as Qualtrics® surveys.
Predictor Variables
For both concurrent relations at each wave of testing, and for longitudinal prediction of semantic knowledge, there were two categories of predictors: measures from the self-derivation through memory integration task and measures from the Woodcock-Johnson III and IV tests of cognitive abilities.
Self-derivation through memory integration
The stimuli were 40 pairs of related facts (hereafter, stem facts) that could be used to self-derive new facts (hereafter, self-derivation facts). All facts were true and based on pilot testing with adults, were deemed unlikely to be familiar to children in the target age range. That is, adult testing demonstrated that the facts were unfamiliar to adults and that both members of the fact pairs were necessary to support high levels of production of the self-derivation facts. Specifically, for all 40 stimulus sets, adult performance was at least two-times as high when both members of the fact pair were presented (2-stem condition) relative to when only one member of the fact pair was presented (1- stem condition). Given these findings with adults, we may logically assume that the facts also would be unfamiliar to children, and that their production of the self-derivation facts would depend on exposure to both members of the fact pairs. Consistent with this assumption, at Wave 1, across the age range and for each nominal age group (8, 9, 10, 11, 12 years), performance in the 2-stem condition was reliably higher than performance in the 1-stem condition [12]. The same effect obtained at Wave 2 [13]. Because only 2-stem performance is indicative of self-derivation through integration, in the present manuscript, all analyses are of performance in the 2-stem condition only; 1-stem condition performance is not included in the present report ([12] for analysis of relative performance in the 1- and 2-stem conditions, and evidence that self-derivation depends on integration of separate yet related facts).
An example stimulus set is provided in Figure 1. The stimuli were obtained from the Bauer Lab Integration and Self-derivation Stimulus (BLISS) bank [29] , from which the stimuli are available upon request: (BLISS bank stimulus numbers S002, S051, S055-057, S066-67, S069, S084, S086, S093, S096-97, S108-111, S113, S126-147). The stimuli also included 24 “filler” facts (BLISS bank stimulus numbers F098-102, F104, F106-107, F109-110, F119-132). Filler facts were structurally similar to the stem facts yet could not be integrated with one another to derive new facts. For purposes of the present report, the purpose of the filler facts was to permit an independent test of fact recall (see below).
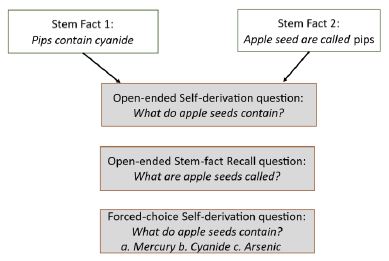
Figure 1: Example stimulus set, including a pair of related stem facts (Stem Facts 1 and 2), example open-ended test questions for self-derivation and stem-fact recall, and example forced-choice test question, with sample options.
Measures of cognitive abilities
Adapted versions of four tests from the Woodcock-Johnson Tests of Cognitive Abilities were used as candidate predictors of measures of domain knowledge: from the Woodcock Johnson III [30], (a) Test 1: Verbal Comprehension, which assesses comprehension of individual words and relations among words across the four subtests of Picture Vocabulary [1a], Synonyms [1b], Antonyms [1c] and Analogy [1d]; and from the Woodcock-Johnson IV [31]; (b) Test 7: Visualization, which is a two-part test of spatial relations, requiring visual-spatial recognition (Spatial Relations [Test 7a]) and mental manipulation of two- and three-dimensional visual representations (Block Rotation [Test 7b]); (c) Test 10: Numbers Reversed, which requires the participant to listen to and then recall a sequence of digits in reverse of the order of presentation. It is considered a measure of working memory; and (d) Test 13, Visual-Auditory Learning, which requires the participant to learn and recall pictographic representations of words. It assesses long-term storage and retrieval as well as associative memory. Verbal Comprehension is considered a measure of semantic memory and thus a “crystalized” ability, whereas Visualization, Numbers Reversed, and Visual-Auditory Learning are considered measures of “fluid” abilities. Due to the online format of data collection, the tests were rendered as Qualtrics® surveys.
Procedure
At each wave of testing, children took part in two sessions conducted and recorded via Zoom. Testing was conducted by one of six female experimenters. Within a wave, children were tested by the same experimenter for both sessions. Children were tested by different experimenters at Waves 1 and 2. The experimenters followed a detailed written protocol. Consistency was assessed by regular viewing of the session recordings by all experimenters. The protocols were the same at Wave 1 and Wave 2. Both within and between sessions, the order of the tasks and tests was the same for all participants. The order of administration was determined based on pilot testing with the goals of minimizing participant burden and maximizing participant engagement. Because the order was fixed, any general or specific carry-over effects are the same for all participants. A schematic of the testing protocol is provided in Figure 2, with Session 1 in the left panel and Session 2 in the right panel.
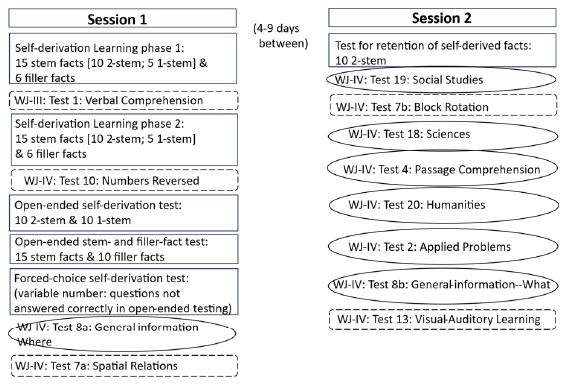
Figure 2: Schematic representation of the testing protocol. Tests in squared boxes are from the self-derivation through memory integration task; tests in rounded rectangles with broken/dashed outlines are measures of cognitive abilities; tests surrounded by ovoids are outcome measures.
Session 1
At each wave, in Session 1, participants engaged in the self- derivation through integration task, three tests of cognitive abilities, and one test of semantic knowledge.
Self-derivation through memory integratiaon
Learning phases. At each wave, in each of two learning phases, children were exposed to 21 novel facts, for a total of 42 facts. Of the 42 facts, 20 were in a 2-stem condition, 10 were in a 1-stem condition, and the remaining 12 were filler facts. Pairs of facts in the 2-stem condition could be integrated such that participants could self-derive new knowledge that had not been directly presented (i.e., self-derivation facts). The 10 facts presented in the 1-stem condition served as a control: each fact was one half of an integrable pair and thus there was no opportunity for integration and self-derivation. As noted above, because only 2-stem performance is indicative of self- derivation through integration, in the present manuscript, all analyses are of performance in the 2-stem condition only; 1-stem condition performance is not included in the report [12,13]. Each integrable fact pair was used approximately equally often in the 2-stem and 1-stem conditions across participants. The 12 filler facts provided the opportunity to examine recall of directly-taught facts, independent of the self-derivation task.
The 42 novel facts were presented across two learning phases. In Learning phase 1, children were presented with 21 facts: the first member of 10 pairs of facts in the 2-stem condition, 5 facts in the 1-stem condition, and 6 filler facts. Children were instructed to pay attention to the facts because they might be asked some questions about them later. Facts were presented individually on the experimenter’s screen, which was shared with the participant via Zoom. The experimenter first read the fact to the child and then the child read it back to the experimenter. After all 21 facts had been displayed and read aloud, children completed the Woodcock Johnson III [30] test of Verbal Comprehension (Test 1; see below for procedure). After the test for verbal comprehension, children engaged in Learning phase 2, during which they were presented with the remaining 21 facts: 10 facts in the 2-stem condition, 5 facts in the 1-stem condition, and 6 filler facts. Participants then completed Woodcock Johnson IV [27,30] Numbers Reversed test (Test 10), as a measure of auditory working memory (see below for procedure).
In total, there were 40 pairs of related stem facts, divided into two sets of 20 pairs of related facts. At each wave, half of the participants were tested on one set of related facts and half on the other. The stem-fact sets were used approximately equally often in each wave of testing, and participants were tested on different sets of facts at each wave. Within a stem-fact set, each stem-fact pair was used in the 2-stem and 1-stem conditions approximately equally often. In the 2-stem condition, each member of the stem-fact pair was presented in Learning phase 1 and Learning phase 2 approximately equally often. Each stem-fact set was presented in one of four different random orders, each used approximately equally often across participants. Participants were pseudo-randomly assigned to one of the four orders, constrained by the need to use each order approximately equally often.
Self-derivation through integration
Test phases. Following the Woodcock Johnson IV Numbers Reversed test (Test 10), children were presented with 20 open-ended questions testing for self-derivation through memory integration: 10 questions on facts from each of the 2-stem and 1-stem conditions. They then were presented with 15 open-ended stem-fact and 10 filler- fact recall questions. After tests for open-ended recall of stem and filler facts, children were tested on any self-derivation questions answered incorrectly in open-ended testing, this time in 3-alternative forced- choice format (see Figure 1 for example open-ended and forced- choice questions). Forced-choice testing of stem and filler facts was not conducted, in consideration of participant burden.
At each wave, the order of presentation of open-ended self- derivation through integration questions was randomized in Qualtrics. As well, the order of forced-choice testing of self-derivation questions not answered correctly in open-ended testing also was randomized in Qualtrics. Open-ended testing for recall of the stem and filler facts was conducted in one of eight pseudo-random orders each of which was used approximately equally often across participants. The constraint was that stem facts from the same fact set were not tested sequentially. Across participants, stem and filler facts were tested approximately equally often. After testing of self-derivation through integration and fact recall, children completed a test of domain knowledge (WJ-IV: Test 8a) and then were administered the third Woodcock-Johnson test: Test 7a: Visualization-Spatial Relations (see below for procedure).
Woodcock-Johnson tests of cognitive abilities
Between Learning phases 1 and 2, children were tested on the WJ- III test Verbal Comprehension (Test 1a-d). Between Learning phase 2 and the tests for self-derivation and fact recall, children were tested on the WJ-IV test of Numbers Reversed test (Test 10). As the final test of the session, children were tested on the WJ-IV test of Visualization- Spatial Relations (Test 7a). The tests were administered following the WJ protocols, with test items displayed on the Zoom screen. Children’s responses were recorded by the experimenter.
Woodcock-Johnson test of semantic knowledge
The pen-ultimate task of Session 1 was a test of academically- related content knowledge WJ-IV: General Information-where (Test 8a). The test was administered following the WJ protocols, with test items displayed on the Zoom screen. Children’s responses were recorded by the experimenter.
Session 2
Session 2 took place approximately 1 week after Session 1. Children were tested for retention of the self-derived facts from Session 1, six tests of academically-related content knowledge, and two tests of cognitive abilities (see Figure 2).
Retention of self-derivation facts
At each wave, children were tested for retention of the self- derivation facts from the 2-stem condition of Session 1. However, because retention of self-derived facts was not a significant predictor of semantic knowledge [12], we did not include measures of retention in analyses.
Woodcock-Johnson tests of semantic knowledge/achievement
Six tests of achievement were administered in the standard order as indicated: Test 19: Social Studies; Test 18: Sciences; Test 4: Passage comprehension; Test 20: Humanities; Test 2: Applied Problems; and Test 8b: General Information-what. All tests were administered following the WJ-IV protocol, with questions displayed on the Zoom screen.
Woodcock-Johnson tests of cognitive abilities
At Session 2, children completed two tests of cognitive abilities (see Figure 2). Between Tests 19 and 18, children completed WJ-IV test of Visualization—Block Rotation (Test 7b). As the final test of the session, children completed the WJ-IV test of Visual-Auditory Learning (Test 13). The tests were administered following the WJ protocols, with test items displayed on the Zoom screen. Children’s responses were recorded by the experimenter.
Scoring and Data Reduction
Scoring was conducted the same way for Waves 1 and 2. For open-ended self-derivation through integration and open-ended recall of stem and filler facts, 1 point was awarded for each correct response, for a total possible of 10 self-derivation facts in the 2-stem condition, 10 self-derivation facts in the 1-stem condition, 15 stem facts (5 stem-fact pairs from the 2-stem condition [10 facts] and 5 facts from the 1-stem condition), and 10 filler facts. Levels of self- derivation performance and of open-ended recall of the stem and filler facts at Session 1 were considered as potential predictors of semantic knowledge (along with measures from the Woodcock-Johnson tests; see below). Self-derivation also was tested in forced-choice. However, because open-ended performance provides stronger evidence of self- derivation, relative to performance that also includes selection of correct responses from among distracters, in subsequent analyses, we focus on open-ended self-derivation performance only ([12], for further justification for focus on open-ended performance only).
The tests of cognitive abilities were scored as per the test protocol and standardized using WJ-III and WJ-IV proprietary software.
Results
The results are presented in four sections, starting with descriptive statistics and zero-order correlations among the variables in Section 1. Section 2 outlines the results of factor analyses to determine the number of latent factors among (a) the outcome measures of domain knowledge; and (b) the predictors of domain knowledge, namely, the (i) tests of cognitive abilities (verbal comprehension, visual-auditory learning, visualization, numbers reversed) and (ii) experimental measures of self-derivation through integration performance, and memory for stem and filler facts. In Section 3 we present the results of tests for concurrent relations among the outcome measures and the predictors at each of Waves 1 and 2. Finally, in Section 4, we present the results of longitudinal prediction of the outcome measures by the predictors from Wave 1 to Wave 2, one year later.
Section 1: Descriptive Statistics and Zero-order Correlations
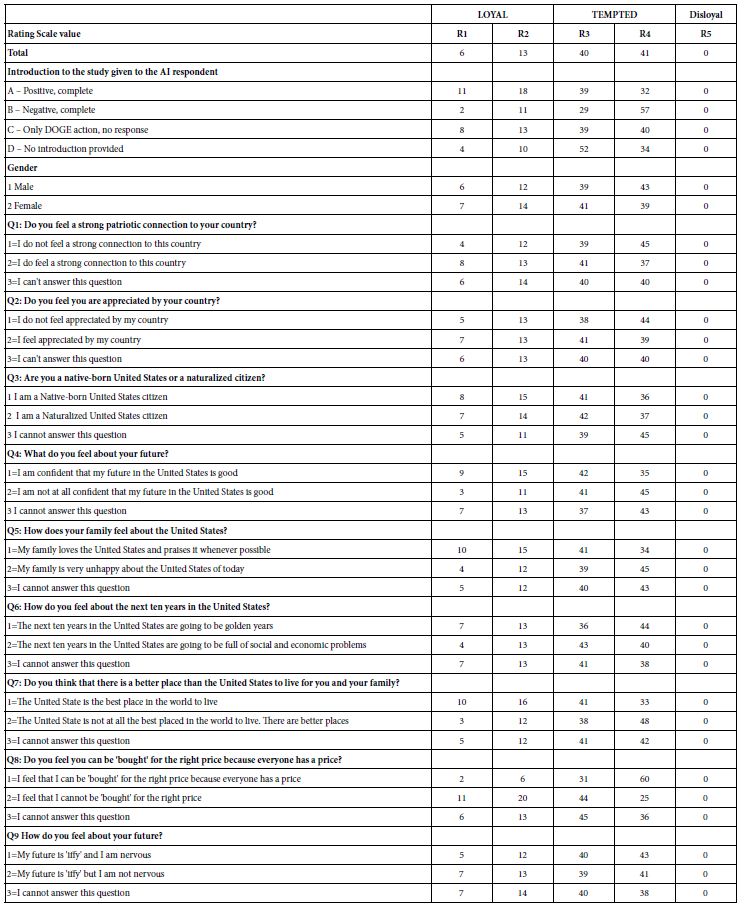
Descriptive statistics for the measures of interest are provided in Table 1, Panels a and b for Wave 1 and Wave 2, respectively. For the outcome measure of domain knowledge, following [13], we report W Scores for each of the six Woodcock-Johnson Tests probing domain knowledge. W Scores represent the child’s performance on the task, based on the average performance of neuro-typical children of their age (in months). For every raw score, there is a W Score that is generated by Woodcock-Johnson Score Reports. W scores are centered on a value of 500, with a typical range on any given task of between 430 and 550. As discussed in [13], W scores represent an equal interval scale across tasks, making them particularly relevant for reporting participants’ actual growth in a measured trait [32]. For all other measures, we report proportion correct out of total trials. Initial skewness and kurtosis measures indicated that the data for all variables were normally distributed, with the exception of the Wave 2 measure of applied problems. One participant performed more than 5 standard deviations below the mean on this assessment. With the data from that participant removed, skewness and kurtosis values indicated normal distribution, as indicated in Table 1. Whether the data from this participant were or were not removed, overall results from subsequent analyses did not change.
Table 1: Descriptive statistics for outcome variables (domain knowledge) and predictor variables (cognitive abilities, self-derivation) at Wave 1 (Panel a) and Wave 2 (Panel b)
|
Category and Measure |
Descriptive Statistics | ||||||
| Category | Measure | N | Model Label | Mean | SD | Skewness |
Kurtosis |
| Panel a: Wave 1 | |||||||
| Domain knowledge | Domain knowledge total |
148 |
Total01 | 3020.42 | 80.74 | -.16 |
.40 |
| General information |
148 |
Gen01 | 503.51 | 14.19 | -.274 |
-.214 |
|
| Passage comprehension |
148 |
PC01 | 499.95 | 14.51 | .022 |
-.154 |
|
| Sciences |
148 |
Sci01 | 501.52 | 14.24 | -.263 |
-.593 |
|
| Applied problems |
148 |
App01 | 509.63 | 19.83 | -.697 |
.958 |
|
| Social studies |
148 |
Soc01 | 505.98 | 17.26 | -.084 |
-.624 |
|
| Humanities |
148 |
Hum01 | 499.83 | 15.75 | .194 |
-.329 |
|
| Cognitive abilities | Verbal comprehension |
143 |
VC01 | 107.90 | 10.755 | .032 |
-.613 |
| Visual-Auditory learning |
147 |
VisAud01 | 105.551 | 13.426 | .007 |
.395 |
|
| Visualization |
148 |
Vis01 | 104.541 | 13.543 | -.029 |
.438 |
|
| Numbers Reversed |
138 |
Num01 | 107.754 | 15.325 | -.524 |
2.151 |
|
| Self-derivation | Open-ended self-derivation |
148 |
SDI01 | .329 | .206 | .354 |
-.423 |
| Stem fact recall |
148 |
Stem01 | .488 | .186 | .015 |
-.376 |
|
| Filler fact recall |
148 |
Fill01 | .567 | .202 | -.328 |
.137 |
|
| Panel b: Wave 2 | |||||||
| Domain knowledge | Domain knowledge total |
148 |
Total02 | 3060.14 | 78.09 | -.21 |
-.28 |
| General information |
148 |
Gen02 | 509.19 | 13.13 | -.075 |
-.158 |
|
| Passage comprehension |
148 |
PC02 | 506.43 | 14.54 | .067 |
.034 |
|
| Sciences |
148 |
Sci02 | 507.80 | 13.25 | -.481 |
-.355 |
|
| Applied problems |
147 |
App02 | 517.57 | 16.69 | -.54 |
.93 |
|
| Social studies |
148 |
Soc02 | 513.35 | 16.54 | -.378 |
-.676 |
|
| Humanities |
148 |
Hum02 | 506.39 | 16.35 | .200 |
-.453 |
|
| Cognitive abilities | Verbal comprehension |
143 |
VC02 | 107.783 | 11.368 | .066 |
.005 |
| Visual-Auditory learning |
144 |
VisAud02 | 113.465 | 12.847 | .64 |
.902 |
|
| Visualization |
148 |
Vis02 | 105.653 | 14.583 | .016 |
.056 |
|
| Numbers Reversed |
143 |
Num02 | 108.329 | 16.369 | -.109 |
-.31 |
|
| Self-derivation | Open-ended self-derivation |
148 |
SDI02 | .388 | .209 | .086 |
-.807 |
| Stem fact recall |
148 |
Stem02 | .533 | .188 | -.225 |
-.449 |
|
| Filler fact recall |
148 |
Fill02 | .592 | .197 | -.313 |
.011 |
|
Note. W Scores are used for all measures of domain knowledge (domain knowledge total, general information, passage comprehension, sciences, applied problems, social studies, humanities); proportion scores are used for measures of self-derivation, including stem-fact and filler-fact recall.
Zero-order correlations among the outcome variables (domain knowledge) and predictor variables (cognitive abilities, self- derivation), within and across waves, are provided in Table 2. Pearson’s product-moment correlations between the categories of predictor variables (cognitive abilities and self-derivation), within and across waves, are provided in Table 3. Pearson’s product-moment correlations within predictor variables (cognitive abilities, self-derivation), within and across waves, are provided in Table 4.
Table 2: Pearson’s product-moment correlations among outcome variables (domain knowledge) and predictor variables (cognitive abilities, self-derivation), within and across waves
|
Predictor Variables |
Age | Measures of Outcome Variables: Domain Knowledge | |||||||||||||
| Age 1 | Total1 | Gen1 | PC1 | Sci1 | App1 | SS1 | Hu1 | Total2 | Gen2 | PC2 | Sci2 | App2 | SS2 |
Hu2 |
|
|
VC1 |
-.38 | .39 | .38 | .37 | .39 | .20** | .28 | .40 | .49 | .46 | .44 | .41 | .30 | .37 | .51 |
| VA1 | .01ns | .42 | .32 | .36 | .35 | .32 | .32 | .47 | .49 | .42 | .40 | .40 | .39 | .35 |
.52 |
|
Vis1 |
-.09ns | .24** | .16* | .26** | .26 | .22** | .09ns | .24** | .31 | .27 | .24** | .23** | .30 | .16ns | .31 |
| NR1 | .09ns | .20* | .17* | .22** | .01ns | .25** | .13ns | .20* | .22** | .19* | .19* | -.01ns | .24** | .18* |
.22* |
|
SDI1 |
.15ns | .59 | .54 | .52 | .45 | .37 | .52 | .60 | .61 | .52 | .50 | .59 | .46 | .51 | .61 |
| Stm1 | .20* | .60 | .55 | .50 | .49 | .42 | .54 | .58 | .62 | .56 | .48 | .55 | .47 | .55 |
.61 |
|
Fill1 |
.16ns | .53 | .42 | .50 | .45 | .35 | .45 | .56 | .57 | .47 | .51 | .56 | .41 | .44 | .58 |
| VC2 | -.20* | .55 | .54 | .57 | .51 | .30 | .40 | .54 | .64 | .61 | .59 | .56 | .43 | .47 |
.65 |
|
VA2 |
.05ns | .42 | .31 | .38 | .36 | .37 | .29 | .43 | .47 | .41 | .38 | .44 | .41 | .30 | .45 |
| Vis2 | 0ns | .30 | .26** | .29 | .30 | .27** | .11ns | .31 | .36 | .32 | .28 | .32 | .32 | .16ns |
.40 |
|
NR2 |
.04ns | .16ns | .14ns | .20* | .02ns | .22* | .10ns | .13ns | .20* | .15ns | .18* | .00ns | .21* | .19* | .16ns |
| SDI2 | .11ns | .61 | .46 | .53 | .56 | .45 | .55 | .52 | .63 | .57 | .49 | .55 | .48 | .56 |
.62 |
|
Stm2 |
.06ns | .55 | .44 | .48 | .52 | .38 | .49 | .48 | .62 | .57 | .51 | .60 | .43 | .50.61 | |
| Fill2 | -.01ns | .52 | .43 | .44 | .48 | .31 | .45 | .54 | .59 | .54 | .47 | .57 | .39 | .48 |
.62 |
Note: Measures ending in “1” were collected at Wave 1; measures ending in “2” were collected at Wave 2. Age = participant age, Total = total domain knowledge, Gen = general information, PC = passage comprehension, Sci = sciences, App = applied problems, SS = social studies, and Hu = humanities. VC = verbal comprehension, VA = visual-auditory learning, Vis = visualization, NR = numbers reversed, SDI = self-derivation, Stm = stem-fact recall, and Fill = filler-fact recall. Unless otherwise indicated, all correlations are significant at p < .001. For other correlations, **= p < .01, * = p < .05, ns = not statistically significant.
Table 3: Pearson’s product-moment correlations between the categories of predictor variables (cognitive abilities, self-derivation), within and across waves
|
Measures of Self-derivation |
Age | Measures of Cognitive Abilities | |||||||
| Age1 | VC1 | Vis-Aud1 | Visual1 | NR1 | VC2 | Vis-Aud2 | Visual2 |
NR2 |
|
|
SDI1 |
.15ns | .36** | .30** | .16* | .13ns | .51** | .35** | .27** | .06ns |
| Stem1 | .20* | .35** | .39** | .18* | .19* | .42** | .37** | .34** |
.22* |
|
Fill1 |
.16ns | .43** | .33** | .25** | .12ns | .49** | .39** | .30** | .15ns |
| SDI2 | .11ns | .41** | .34** | .18* | .14ns | .48** | .33** | .26** |
.14ns |
|
Stem2 |
.06ns | .45** | .44** | .23** | .09ns | .52** | .42** | .30** | .12ns |
| Fill2 | -.01ns | .52** | .42** | .25** | .13ns | .56** | .43** | .31** |
.11ns |
Note: Measures ending in “1” were collected at Wave 1; measures ending in “2” were collected at Wave 2. Age = participant age, VC = verbal comprehension, Vis-Aud = visual-auditory learning, Visual = visualization, NR = numbers reversed, SDI = self-derivation, stem = stem-fact recall, and Fill = filler-fact recall. ** = p < .001, * = p < .05, ns = not statistically significant.
Table 4: Pearson’s product-moment correlations within predictor variables (cognitive abilities, self-derivation), within and across waves
|
Measures of Predictor Variables at Wave 2 |
Measures of Predictor Variables at Wave 1 | ||||||
| VC1 | Vis-Aud1 | Visual1 | NR1 | SDI1 | Stem1 |
Filler1 |
|
|
VC2 |
.801** | .48** | .42** | .15ns |
(See Table 3 for values in this quadrant) |
||
| Vis-Aud2 | .35** | .582** | .32** |
.14ns |
|||
|
Visual2 |
.30** | .44** | .71** | .24** | |||
| NR2 | .12ns | .19* | .21* |
.66** |
|||
|
SDI2 |
(See Table 3 for values in this quadrant) |
.537** | .65** | .50** | |||
| Stem2 | .58** | .632** |
.60** |
||||
|
Filler2 |
.56** |
.65** |
.556** |
||||
Note: Measures ending in “1” were collected at Wave 1; measures ending in “2” were collected at Wave 2. Age = participant age, VC = verbal comprehension, Vis-Aud = visual-auditory learning, Visual = visualization, NR = numbers reversed, SDI = self-derivation, Stem = stem-fact recall, and Fill = filler-fact recall. ** = p < .001, * = p < .05, ns = not statistically significant.
Section 2: Factor Analyses
We conducted factor analyses to determine the number of latent factors among (a) the outcome measures of domain knowledge; and (b) the predictors of domain knowledge, namely, the (i) tests of cognitive abilities (verbal comprehension, visual-auditory learning, visualization, numbers reversed) and (ii) experimental measures of self-derivation through integration performance, and memory for stem and filler facts.
Outcome measures: Domain knowledge
Exploratory factor analysis (EFA) reported in [13] indicated that the six domains of knowledge formed one latent factor, thereafter referred to as knowledge. The specific procedures used to arrive at this solution are detailed in [13].
Predictors
For the Wave 1 variables, we used EFA to determine the number of latent factors formed by the tests of cognitive abilities (verbal comprehension, visual-auditory learning, visualization, numbers reversed) and the measures from the self-derivation through integration paradigm (open-ended self-derivation, stem-fact recall, filler-fact recall). The analysis was conducted using JASP software with the following settings: number of factors based on parallel analysis of factors, using the minimum residual estimation method, oblique rotation (promax), and with a factor loading cut off of .4. For Wave 2, given the absence of a theoretical expectation of a different factor structure, we conducted a confirmatory factor analysis (CFA) on the new data, using the factor structure discovered at Wave 1.
For the Wave 1 assessments, a Kaiser-Meyer-Olkin test indicated that the measures were suitable for factor analysis, with an overall MSA =.776. Bartlett’s test was significant, X2(21) = 322.33, p < .001, indicating that the data are a good fit to the factor analysis. As depicted in Table 5, two factors emerged: Factor 1 was comprised of the measures of stem-fact recall, open-ended self-derivation through integration, and filler-fact recall (hereafter self-derivation and fact recall). Factor 2 was comprised of the measures of the cognitive abilities of visual-auditory learning, verbal comprehension, and visualization (hereafter verbal and spatial abilities). The test of working memory (WJ-IV, Numbers Reversed) did not load on either factor. For the confirmatory factor analysis of the Wave 2 data, a Kaiser-Meyer- Olkin test indicated that the measures were suitable for factor analysis, with an overall MSA = .827. Bartlett’s test was significant, X2(15) = 338.269, p < .001, indicating good model fit. RMSEA = .069, indicating acceptable model fit. The parameter estimates are provided in Table 6.
Table 5: Results of exploratory factor analysis (EFA) for the predictor variables (self-derivation through integration, cognitive abilities) at Wave 1
|
Predictor Variables |
Factor Structure | ||
| Factor 1
(self-derivation and fact recall) |
Factor 2
(verbal and spatial abilities) |
Uniqueness |
|
| Stem-fact recall |
0.906 |
0.215 |
|
| Self-derivation |
0.877 |
0.307 |
|
| Filler-fact recall |
0.642 |
0.468 |
|
| Visual-Auditory Learning |
|
0.703 |
0.484 |
| VerbalComprehension |
|
0.618 |
0.531 |
| Visualization |
|
0.614 |
0.687 |
| Numbers Reversed |
|
0.919 |
|
Note. Applied rotation method is promax.
Table 6: Parameter estimates for confirmatory factor analysis (CFA) for the predictor variables (self-derivation and fact recall, verbal and spatial abilities) at Wave 2: Factor loadings (Panel a) and residual variances (Panel b)
|
Factor and Indicator |
Parameter Estimates | ||||||||
|
95% Confidence Interval |
|||||||||
|
Factor |
Indicator | Estimate | Std. Error | z-value | p | Lower |
Upper |
||
| Panel a: Factor loadings | |||||||||
| Self-derivation & fact recall | Self-derivation |
0.171 |
0.015 | 11.677 | < .001 | 0.142 |
0.2 |
||
| Filler-fact recall |
0.143 |
0.015 | 9.695 | < .001 | 0.114 |
0.172 |
|||
| Stem-fact recall |
0.169 |
0.013 | 13.211 | < .001 | 0.144 |
0.194 |
|||
| Verbal & spatial abilities | Verbal comprehension |
8.543 |
0.955 | 8.942 | < .001 | 6.671 |
10.416 |
||
| Visualization |
7.008 |
1.297 | 5.404 | < .001 | 4.466 |
9.55 |
|||
| Visual-Auditory Learning |
7.632 |
1.117 | 6.832 | < .001 | 5.443 |
9.822 |
|||
| Panel b: Residual variances | |||||||||
| Self-derivation & fact recall | Self-derivation |
0.14 |
.002 | 6.233 | < .001 | 0.010 |
0.018 |
||
| Filler-fact recall |
0.018 |
.003 | 7.134 | < .001 | 0.013 |
0.023 |
|||
| Stem-fact recall |
0.007 |
.002 | 3.835 | < .001 | 0.003 |
0.010 |
|||
| Verbal & spatial abilities | Verbal comprehension |
53.965 |
11.357 | 4.752 | < .001 | 31.705 |
76.224 |
||
| Visualization |
162.122 |
20.923 | 7.749 | < .001 | 121.114 |
203.131 |
|||
| Visual-Auditory Learning |
105.082 |
15.182 | 6.921 | < .001 | 75.325 |
134.838 |
|||
Section 3: Concurrent Relations among Outcome Measures and Predictors
To examine concurrent relations among the outcome measures of knowledge and the predictors of self-derivation and fact recall and verbal and spatial abilities, we conducted Structural Equation Modeling (SEM) at each of Waves 1 and 2. At each wave, we used the two latent structures discovered using factor analyses (self- derivation and fact recall, verbal and spatial abilities) as predictors of the latent outcome variable of knowledge. We did not include the WJ-IV measure of Numbers Reversed (working memory) because, as discussed in Bauer et al. (2025), SEM modeling that included working memory was less than an ideal fit; the model without working memory was a better fitting model. Accordingly, in subsequent analyses, we used the model with only the latent constructs. At both waves, Chi-square goodness of fit tests indicated good model fit: X2(12) = 12.164, p = .433, X2(12) = 17.687, p = .126, for Waves 1 and 2, respectively. Model statistics are provided in Table 7, Panels a and b for Wave 1, and Panels c and d for Wave 2. Semi-partial R-squared values indicating the proportion of variance explained by each factor are provided in Table 8, Panel a.
Table 7: Parameter estimates for Structural Equation Modeling (SEM) of the outcome measure of knowledge with the predictor variables (verbal and spatial abilities, self-derivation and fact recall) at Wave 1: Factor loadings (Panel a) and regression coefficients (Panel b), and Wave 2: Factor loadings (Panel c) and regression coefficients (Panel d)
|
Factor and Indicator |
Parameter Estimates |
||||||
| Panel a: Wave 1 Factor loadings | |||||||
|
95% Confidence Interval |
|||||||
| Latent | Indicator |
Estimate |
Std. Error | z-value | p | Lower |
Upper |
| Verbal & spatial abilities1 | Verbal comprehension1 |
1.000 |
.000 | 1.000 |
1.000 |
||
| Visualization1 |
0.873 |
0.182 | 4.808 | < .001 | 0.517 |
1.229 |
|
| Visual-Auditory Learning1 |
1.277 |
0.213 | 6.005 | < .001 | 0.860 |
1.694 |
|
| Self-derivation & fact recall1 | Self-derivation1 |
1.000 |
.000 | 1.000 |
1.000 |
||
| Filler-fact recall1 |
0.864 |
0.091 | 9.538 | < .001 | 0.687 |
1.042 |
|
| Stem-fact recall1 |
0.959 |
0.082 | 11.650 | < .001 | 0.798 |
1.121 |
|
| Panel b: Wave 1 Regression coefficients | |||||||
|
95% Confidence ICnterval |
|||||||
| Predictor | Outcome |
Estimate |
Std. Error | z-value | p | Lower |
Upper |
| Verbal & spatial abilities1 | Knowledge1 |
2.235 |
1.063 | 2.103 | < .036 | 0.152 |
4.319 |
| Self-derivation & fact recall1 | Knowledge1 |
273.283 |
45.282 | 6.035 | < .001 | 184.533 |
362.034 |
| Panel c: Wave 2 Factor loadings | |||||||
|
95% Confidence Interval |
|||||||
| Latent | Indicator |
Estimate |
Std. Error | z-value | p | Lower |
Upper |
| Verbal & spatial abilities2 | Verbal comprehension2 |
1.000 |
.000 | 1.000 |
1.000 |
||
| Visualization2 |
0.773 |
0.150 | 5.160 | < .001 | 0.479 |
1.066 |
|
| Visual Auditory Learning2 |
0.852 |
0.133 | 6.400 | < .001 | 0.591 |
1.112 |
|
| Self-derivation & fact recall2 | Open-ended self-derivation2 |
1.000 |
.000 | 1.000 |
1.000 |
||
| Filler-fact recall2 |
0.827 |
0.085 | 9.778 | < .001 | 0.661 |
0.993 |
|
| Stem-fact recall2 |
0.948 |
0.078 | 12.188 | < .001 | 0.796 |
1.101 |
|
| Panel d: Wave 2 Regression coefficients | |||||||
|
95% Confidence Interval |
|||||||
| Predictor | Outcome |
Estimate |
Std. Error | z-value | p | Lower |
Upper |
| Verbal & spatial abilities2 | Knowledge2 |
5.284 |
1.609 | 3.284 | < .001 | 2.130 |
8.437 |
| Self-derivation & fact recall2 | Knowledge2 |
121.647 |
69.387 | 1.753 | 0.080 | -14.349 |
257.643 |
Note: Measures ending in “1” were collected at Wave 1; measures ending in “2” were collected at Wave 2.
At Wave 1, both self-derivation and fact recall and verbal and spatial abilities contributed significant unique variance in predicting knowledge: 33.3% and 4.5%, respectively (Table 8). Shared variance was 18.4%, bringing combined variance explained to R2 = .56. At Wave 2, verbal and spatial abilities once again contributed significant unique variance in knowledge (35.4%). However, the latent factor of self- derivation and fact recall merely approached significance, with 7.4% variance accounted for. The amount of shared variance was 27.3%, bringing combined variance explained to R2 = .70. Discussion of the most likely explanations for this pattern is reserved for the Discussion.
Table 8: Beta weights and semi-partial R2 values for concurrent predictors of knowledge at each of Waves 1 and 2 (Panel a) and longitudinal predictors of knowledge from Wave 1 to Wave 2 (Panel b)
|
Wave |
Beta Weights | Variance Explained |
Type of Variance |
||||
| Wave |
ß(VVA) |
ß(SDI) | r(VVA,SDI) | R2 (total) | Unique VVA% | Unique SDI% |
Shared% |
| Panel a: Concurrent prediction | |||||||
| Wave 1 |
.212 |
.577 | .754 | .562 | 4.5% | 33.3% |
18.4% |
| Wave 2 |
.595 |
.272 | .843 | .701 | 35.4% | 7.4% |
27.3 |
| Panel b: Longitudinal prediction | |||||||
| Wave 1 to Wave 2 |
.379 |
.507 | .635 | .625 | 9.3% | 15.8% |
37.4% |
Note: VVA = verbal and visual abilities, SDI = self-derivation and fact recall.
Section 4: Longitudinal Relations among Outcome Measures and Predictors
To examine longitudinal relations among the outcome measures of knowledge at Wave 2 and the predictors of self-derivation and fact recall and verbal and spatial abilities at Wave 1, we conducted SEM. We used the two latent structures of verbal and spatial abilities and self-derivation and fact recall from Wave 1 as predictors of the latent outcome variable of knowledge at Wave 2. A Chi-square goodness of fit test indicated good model fit: X2(12) = 12.088, p = .439. Model statistics are provided in Table 9. As reflected in Table 8, Panel b, both self-derivation and fact recall and verbal and spatial abilities at Wave 1 were significant predictors of knowledge one-year later, at Wave 2, explaining 15.8% and 9.3%, respectively. Shared variance was 37.4%, bringing the combined variance explained to R2 = .63.
Table 9: Parameter estimates for Structural Equation Modeling (SEM) of the outcome measure of knowledge at Wave 2 with the predictor variables (verbal and spatial abilities, self-derivation and fact recall) from Wave 1: Factor loadings (Panel a) and regression coefficients (Panel b)
|
Factor and Indicator |
Parameter Estimates |
||||||
| Panel a: Wave 1 to Wave 2 factor loadings | |||||||
|
95% Confidence Interval |
|||||||
| Latent | Indicator |
Estimate |
Std. Error | z-value | p | Lower |
Upper |
| Verbal & spatial abilities1 | Verbal comprehension1 |
1.000 |
.000 | 1.000 |
1.000 |
||
| Visualization1 |
0.865 |
0.174 | 4.967 | < .001 | 0.524 |
1.206 |
|
| Visual Auditory Learning1 |
1.251 |
0.193 | 6.486 | < .001 | 0.873 |
1.629 |
|
| Self-derivation & fact recall1 | Self-derivation1 |
1.000 |
.000 | 1.000 |
1.000 |
||
| Filler-fact recall1 |
0.863 |
0.090 | 9.608 | < .001 | 0.687 |
1.039 |
|
| Stem-fact recall1 |
0.949 |
0.081 | 11.675 | < .001 | 0.790 |
1.109 |
|
| Panel b: Wave 1 to Wave 2 Regression coefficients | |||||||
|
95% Confidence Interval |
|||||||
| Predictor | Outcome |
Estimate |
Std. Error | z-value | p | Lower |
Upper |
| Verbal & spatial abilities1 | Knowledge2 |
3.829 |
1.014 | 3.774 | < .001 | 1.841 |
5.817 |
| Self-derivation & fact recall1 | Knowledge2 |
231.203 |
40.682 | 5.683 | < .001 | 151.467 |
310.939 |
Note: Measures ending in “1” were collected at Wave 1; measures ending in “2” were collected at Wave 2.
Discussion
The major purpose of the present research was to determine whether self-derivation through memory integration makes unique contributions to prediction of domain knowledge and thus can be considered a mechanism of semantic knowledge development in childhood. The question was tested in a sample of 148 children, ages 8 to 12 years at the time of enrollment. The outcome measure of semantic knowledge was based on six domains that were treated as one latent factor, knowledge: applied problems (math), humanities, sciences, social studies, general knowledge of what and where, and passage comprehension. The predictors were (a) tests of self- derivation through memory integration and memory for directly taught facts (stem facts and filler facts); and (b) measures of four cognitive abilities, namely, verbal comprehension, visual-auditory learning, visualization, and working memory. The predictors formed two latent factors: self-derivation and fact recall and verbal and spatial abilities. Working memory was not included in either factor. Children participated in two waves of testing, separated by one year.
In prior research on the same sample, self-derivation through memory integration was found to predict semantic knowledge for five of the six individual domains assessed; the exception was the domain of sciences (Bauer et al., 2024). In a subsequent test of longitudinal relations, the six individual domains were treated as one latent factor, knowledge. Self-derivation through integration predicted knowledge both concurrently, at each of Waves 1 and 2, and longitudinally over one year [13]. In a separate evaluation, measures of memory for directly taught facts (stem and filler facts) were found to be both concurrently related and longitudinally predictive of self-derivation, whereas the measures of cognitive abilities were not [8]. Based on this pattern of findings, in the present research, we expected to observe (a) separate latent factors for self-derivation and fact recall and visual and spatial abilities; and (b) that both latent factors would be predictive of knowledge, concurrently and over one year.
Consistent with expectations, at Wave 1, the latent constructs of self-derivation and fact recall and verbal and spatial abilities both were uniquely predictive of semantic knowledge. The amount of unique and shared variance in semantic knowledge explained by the factors was 56%. At Wave 2, only verbal and spatial abilities was uniquely predictive. Self-derivation and fact recall predicted a nonsignificant 7.4% of the variance in semantic knowledge. Considering the amount of unique and shared variance, the total variance explained by the factors was 70%. Notably, semantic knowledge at Wave 2 was predicted by Wave 1 self-derivation and fact recall and Wave 1 verbal and spatial abilities. Both latent factors made unique contributions to prediction of semantic knowledge at Wave 2. The amount of unique and shared variance in semantic knowledge explained by the factors was 63%.
Overall, the findings of the present research were as expected, with the exception of the concurrent test at Wave 2, in which only verbal and spatial abilities emerged as a statistically significant predictor of semantic knowledge; self-derivation and fact recall only approached significance. We attribute this pattern to the strong correlations among the various Woodcock-Johnson tests at Wave 2 (see Table 2), and to the nominal increase in shared variance of the factors at Wave 2 (27.3%), relative to Wave 1 (18.4%; see Table 8). The result was relatively less variance to be explained by the experimental measures of self- derivation. The pattern of relatively stronger correlations among the Woodcock-Johnson tests at Wave 2, relative to Wave 1 was especially apparent for the measure of verbal comprehension: at Wave 1, the correlations with the individual semantic domains ranged from rs = .20-.40, whereas at Wave 2, they ranged from rs = .43-.65. This finding is understandable in light of the fact that the Woodcock-Johnson tests were essentially identical at the two waves: the same questions were asked in the same format. The only differences were in one or possibly two additional questions posed to establish ceiling performance. Given this feature of the instrument, there was substantial shared task variance at Wave 2, likely inflating the proportion of variance in semantic knowledge accounted for by verbal and spatial abilities. In contrast, whereas the format of the tests for self-derivation and fact recall were the same at both waves, the specific items were unique at each wave.
An unexpected—yet explicable—finding in the present research was of the substantial amount of variance in longitudinal prediction shared by the latent factors of self-derivation and fact recall and verbal and spatial abilities. Whereas each factor contributed unique variance, the amount of shared variance in knowledge explained approached 40%. We interpret this pattern as a reflection of the tight relation between existing knowledge and the ability to add to it. In the present research, existing knowledge was reflected in the latent construct of verbal and spatial abilities, which included a measure often used as a proxy for existing knowledge, namely verbal comprehension. The ability to add new knowledge was reflected in the latent construct of self-derivation and fact recall, which included both memory for directly taught facts and the productive process of generating new knowledge from them. Critically, each individual construct at Wave 1 was a unique predictor of semantic knowledge at Wave 2, over and above the variance they shared. Self-derivation and fact recall accounted for a nominally greater proportion of variance relative to verbal and spatial abilities (15.8% and 9.3%, respectively)
The present research makes a substantial contribution to the literature in establishing self-derivation through memory integration as a unique predictor of semantic knowledge, at least among children ages 8 to 12 years. It is not without limitations, however. One limitation is that, based on the present research, it is not known whether the same pattern of findings would be apparent among children younger than age 8 years or among participants older than 12 years. The structure of cognitive abilities changes across the late elementary school years, with increasing differentiation of cognitive abilities from one another [33]. As such, different patterns may be expected at younger and older ages. It will be left to future research to address this possibility.
A second potential limitation is that the children were tested online, as opposed to in person. The online format may have resulted in lower performance than might be expected from in-person testing. Importantly, given that all tasks and all participants were tested in the same way, any general or specific effects of online testing would be equally distributed over the entire battery and sample.
A third potential limitation of the present research is that only one productive process was tested as a predictor of semantic knowledge. As noted earlier, self-derivation through memory integration is but one among many productive processes, including (but not limited to) analogy, induction, and associative and transitive inference [1,4, 34]. It is possible that one or more of these processes also may account for unique variance in semantic knowledge. This question has yet to be tested empirically in children. Among adults, self-derivation through integration has been found to be uniquely related to academic achievement relative to other paradigms that test productive processes [35]. Importantly, finding that other productive processes also predict semantic knowledge would not undermine the importance of self- derivation through memory integration as a mechanism of semantic knowledge development. Rather, it would establish a larger suite of productive processes on which knowledge expansion depends.
In conclusion, in the present research, we tested self-derivation through memory integration along with measures of other cognitive abilities, as concurrent and longitudinal predictors of semantic knowledge. Both factors were found to be concurrent predictors at Wave 1. Critically, both factors as measured at Wave 1 predicted semantic knowledge at Wave 2. The work thus establishes self- derivation through memory integration as a unique longitudinal predictor of semantic knowledge in childhood. As such, it can be considered a mechanism of semantic knowledge development.
Acknowledgement
Support for this research was provided by NICHD R01 HD094716 to Patricia J. Bauer. The authors also thank Britney Del Solar, Jessica Dugan, Melanie Hanft, and Alissa Miller, for their help with data collection and reduction, and other members of the Memory at Emory laboratory group for their help at various stages of this research, and the children and families who so generously gave of their time to take part in this research.
References
- Richland, L. E., Morrison, R. G., & Holyoak, K. J. (2006). Children’s development of analogical reasoning: Insights from scene analogy problems. Journal of Experimental Child Psychology;94(3): 249–273.
- Bryant, E., & Trabasso, T. (1971). Transitive inferences and memory in young children. Nature, 232(5311):456–458. [crossref]
- Schlichting, M. L., & Preston, A. R. (2015). Memory integration: Neural mechanisms and implications for behavior. Current Opinion in Behavioral Sciences, 1:1-8. [crossref]
- Schulz, E., Goodman, N. D., Tenenbaum, J. B., & Jenkins, A. C. (2008). Going beyond the evidence: Abstract laws and preschoolers’ responses to anomalous data. Cognition, 109(2):211-223. [crossref]
- Goswami, (2011). Inductive and deductive reasoning. In U. Goswami (Ed.), Childhood cognitive development (pp. 399–419). Oxford, UK: Wiley–Blackwell.
- Siegler, S. (1989). Mechanisms of cognitive development. Annual Review of Psychology, 40:353–379. [crossref]
- Bauer, P.J. (2021). We know more than we ever learned: Processes involved in accumulation of world knowledge. Child Development Perspectives, 15(4), 220-227. [crossref]
- Bauer, J., Lee, K. A., Dugan, J. A., & Cronin-Golomb, L. M. (2025). Longitudinal predictors of self-derivation through memory integration—A mechanism of knowledge accumulation. Journal of Experimental Child Psychology. Advanced online publication. doi.org/10.1016/j.jecp.2024.106120
- Bauer, J., & Jackson, F.L. (2015). Semantic elaboration: ERPs reveal rapid transition from novel to known. Journal of Experimental Psychology: Learning, Memory, and Cognition, 41(1):271-282.[crossref]
- Esposito, G., & Bauer, P. J. (2022). Determinants of elementary-school academic achievement: Component cognitive abilities and memory integration. Child Development, 93(6):1777-1792.[crossref]
- Esposito, G., & Bauer, P. J. (2024). Self-derivation through memory integration: A longitudinal examination of performance and relations with academic achievement in elementary classrooms. Cognitive Development, 69:101416.[crossref]
- Bauer, J., Dugan, J. A., Cronin-Golomb, L. M., Lee, K. A., Del Solar, B., et al.(2024). Development of self-derivation through memory integration and relations with world knowledge. Memory, 32(8), 981-995.[crossref]
- Cronin-Golomb, M., & Bauer, P. J. (in press). Longitudinal relations between self-derivation and semantic knowledge growth. Journal of Experimental Child Psychology.[crossref]
- Shing, Y. L., Finke, C., Hoffmann, M., Pajkert, A., Heekeren, H. R., & Ploner, C. J. (2019). Integrating across memory episodes: Developmental trends. PLOS ONE, 14(4), [crossref]
- Bauer J., & Larkina, M. (2017). Realizing relevance: The influence of domain- specific information on generation of new knowledge through integration in 4- to 8-year-old children. Child Development, 88:247-262. [crossref]
- Bauer, J., & San Souci, P. (2010). Going beyond the facts: Young children extend knowledge by integrating episodes. Journal of Experimental Child Psychology, 107(4):452-465.[crossref]
- Esposito, A. G., & Bauer, P. J. (2017). Going beyond the lesson: Self-generating new factual knowledge in the classroom. Journal of Experimental Child Psychology, 153:110-125.
- Miller-Goldwater, E., Williams, B. M., Hanft, M. H., & Bauer, P. J. (2024). Contributions of shared book reading to children’s learning of new semantic facts through memory integration. Early Childhood Research Quarterly; 68:99-111. [crossref]
- Cronin-Golomb, M., Pejic, J., Miller-Goldwater, H. E., & Bauer, P. J. (2024). Factors affecting children’s explicit learning and productive memory processes in the context of virtual museums. Cognitive Development; 71: 101454.[crossref]
- Wilson, J. T., & Bauer, P. J. (2024). Generative and active engagement in learning neuroscience: A comparison of self-derivation and rephrase. Cognition, 245:105709. [crossref]
- Varga, L., & Bauer, P. J. (2017). Young adults self-derive and retain new factual knowledge through memory integration. Memory & Cognition, 45:1014-1027.
- Gathercole, E., Pickering, S. J., Knight, C., & Stegmann, Z. (2003). Working memory skills and educational attainment: Evidence from national curriculum assessments at 7 and 14 years of age. Applied Cognitive Psychology, 18:1-16.
- Serpell, N., & Esposito, A. G. (2016). Development of executive functions: Implications for educational policy and practice. Policy Insights from the Behavioral and Brain Sciences, 3:203–210.
- Alloway, P., & Alloway, R. G. (2010). Investigating the predictive roles of working memory and IQ in academic attainment. Journal of Experimental Child Psychology, 106(1):20-29.[crossref]
- Bull, , Espy, K. A., & Wiebe, S. A. (2008). Short-term memory, working memory, and executive functioning in preschoolers: Longitudinal predictors of mathematical achievement at age 7 years. Developmental Neuropsychology, 33(3):205–228.[crossref]
- Fanari, R., Meloni, C., and Massidda, D. (2019). Visual and spatial working memory abilities predict early math skills: a longitudinal study. Frontiers in Psychology; 10:2460. [crossref]
- Schrank, A., McGrew, K.S., & Mather, N. (2014). Woodcock-Johnson IV Tests of Cognitive Abilities. Rolling Meadows, IL: Riverside.
- Terman, M., Kohs, S. C., Chamberlain, M. B., Anderson, M., & Henry, B. (1918). The vocabulary test as a measure of intelligence. Journal of Educational Psychology, 9(8):452–466.
- Bauer, P. J. (06 May 2020). Bauer Lab Integration and Self-derivation Stimulus (BLISS) bank. PsyArXiv. Doi: 10.31234/osf.io/rv9n7
- Woodcock, W., McGrew, K. S., & Mather, N. (2001). Woodcock-Johnson III Tests of Cognitive Abilities. Itasca, IL: Riverside.
- Schrank, F. A., & Wendling, B. J. (2018). The Woodcock–Johnson IV: Tests of cognitive abilities, tests of oral language, tests of Contemporary intellectual assessment: Theories, tests, and issues, 4th ed (pp. 383–451). The Guilford Press.
- Jaffe, (2009). Development, Interpretation, and Application of the W Score and the Relative Proficiency Index (11; Woodcock-Johnson III Assessment Service). Riverside Publishing.
- Mungas, , Widaman, K., Zelazo, P. D., Tulsky, D., Heaton, R. K., et al. (2013). NIH Toolbox Cognition Battery (CB): Factor structure for 3- to 15-year-olds. Monographs of the Society for Research in Child Development, 78:103–118. [crossref]
- Smith, C., & Squire, L. R. (2005). Declarative memory, awareness, and transitive The Journal of Neuroscience, 25(44):10138-10146. [crossref]
- Varga, N. L., Gaugler, T., & Talarico, J. (2019). Are mnemonic failures and benefits two sides of the same coin?: Investigating the real-world consequences of individual differences in memory integration. Memory & Cognition, 47(3):496-510.[crossref]