DOI: 10.31038/IDT.2022311
Abstract
As the SARS-CoV-2 pandemic is nearing its eventual end we focus on what we believe are two key omissions from the mainstream scientific literature and which have significant implications for how mankind manages the next global pandemic. We therefore review data, observations, analyses and conclusions from our series of papers published through 2020 and 2021 on its likely cometary origin and global spread. We also revisit our long held understanding of the superior effectiveness of intra-nasal vaccines against respiratory tract pathogens that involve induction of dimeric secretory IgA antibodies. While these two oversights seem disparate, together they provide us with new insights into our collective awareness of how we might view and address the next global pandemic. We begin with our hypothesis of the likely cometary origin of the SARS-CoV-2 virus via a bolide strike in the stratosphere on the night of October 11 2019 on the 40o N line over Jilin in NE China. Further global spread most likely occurred via prevailing wind systems transporting both the pristine cometary virus followed by continuing strikes from the same primary source as well as prior human-passaged virus transmitted by person to person spread and through contaminated dust in global wind systems. We also include a discussion of our prior work on data relating to vaccine protective efficacy. Finally we review the totality of evidence concerning the likely origin and global spread of the predominant variants of the virus ‘Omicron’ (+Delta mix?) from early to mid-December 2021 and extending into the first week January 2022. We describe the striking data showing the large numbers of infectious cases per day and outline the scale of what appears to be a global pandemic phenomenon, the causes of which are unclear and not completely understood. Firstly, these essentially simultaneous and sudden global-wide epidemic COVID-19 out breaks, appear to be largely correlated with events external to the Earth, probably causing globally correlated precipitation events. They appear related broadly to “Space Weather” events that render the Earth vulnerable to cosmic pandemic pathogen attack particularly during times of the minima of the Sunspot Solar Cycle which we are now currently passing through. Secondly, we argue that these sudden global-wide epidemic outbreaks of COVID-19 are specifically largely influenced by global wind transport and deposition mechanisms, the physics of which we need to further explore and comprehend. We conclude on an optimistic note for mankind. Given our prior knowledge of the effectiveness against respiratory tract pathogens of mucosal immunity involving induction of dimeric secretory IgA antibodies, we consider that the recently published intra-nasal vaccine data from laboratories based at the University of California, San Francisco and, independently at Yale University. These latter studies hold out great promise for the future development of both pan-specific and specific immunity against future pandemics caused by suddenly emergent respiratory pathogens, whether viral, bacterial or fungal.
Introduction
The authors encompass a multi-disciplinary team across the scientific disciplines of Biology, Medicine and Physics in the broadest meaning of those categories of scientific understanding. We follow in the footsteps of the prior foundation studies published in many key historical works on Astronomy, Astrophysics and Astrobiology incorporating references to many peer reviewed papers (many in the journal Nature) by Fred Hoyle and N. Chandra Wickramasinghe [1-8]. Many authors on the current list of co-authors have made significant contributions to recent publications on diverse related matters, with some of these presciently dating just prior to the emergence of the COVID-19 pandemic [9-13]. This previous experience has heightened our analytical ability to scientifically track and plausibly explain the cometary origin and global spread of COVID-19. A number of reviews of the relevant datasets and conclusions therefore have already been published through 2020 and 2021 [14-16], including a compendium of chapters on ‘Cosmic Genetic Evolution’ all of which places the COVID-19 pandemic in its appropriate cosmic perspective [17]. Thus, a number of papers published by us since February 14 2020 review the data supporting our first claims of COVID-19’s putative meteorite origins over China, following a cometary bolide strike on the 40o N Latitude line over Jilin, NE China on the night of October 11, 2019 [18]. We note here that the causative virus-carrying bolide may not have arrived at the top of the Earth’s atmosphere as a cohesive body, but as an aggregation of dust particles, with individual radii of the order of micrometres. It is well known that of the order of 100 tonnes of such material are incident on the upper atmosphere daily. Approximately two thirds of micron sized micrometeorites can plausibly be assumed to be of cometary origin. Modelling of their dynamics in the atmosphere shows that a significant fraction of these particles reach the surface of the Earth without experiencing destructive heating [19].
Our subsequent publications in the early weeks of the pandemic focused on the relative lack of evidence for person-to person transmission as the primary infection mechanism for COVID-19 [20]. Indeed, detailed analyses of the active region-wide epidemic episodes around the globe during 2020 and 2021 occurred initially (late 2019 to March through April 2020) mainly on the 30-50o N latitude band with limited outbreaks taking place outside this band [21]. We developed explanations of the unfolding data relating to the pandemic as it engulfed the world from the later months of 2019. In our modelling we took full account of genetic, immunologic and epidemiologic evidence, as well as the role of geophysical and atmospheric processes including a possible continuation of a space input of the virus.
In summary, our main analyses were as follows:
a. The genetic analysis of the SARS-CoV-2 viral genomes and the deaminase-mediated haplotype variation and adaptation strategy of the coronavirus as it navigated infections in different susceptible/vulnerable human hosts and genetic backgrounds first in China then Spain, France and New York with eventual infection documented in Australia from January through to September 2020 [15,18,20-23].
b. The immunologic analysis of the host-parasite relationship and vaccine efficacy of systemic versus mucosal-local antigen routes of immunization [22-24].
c. The epidemiologic analyses of both the temporal order of epidemics and their global location, including the role of prevailing winds systems, remote outpost strikes (O’Higgins Chilean Army outpost in Antarctica), sudden island strikes and strikes on ships at sea [15,16,25,26].
d. The geophysics and atmospheric physics of the major convection cells plus jet streams sweeping up and depositing virus-bearing dust, including human-passaged aerosol transferring them in the northern hemisphere with limited connection to the south [21,27-29]. Global connectivity is effective on the 10-day time-scale. The evidence of long-distance tropospheric transportation in the Northern Hemisphere is provided by the COVID-19 genomic sequence data from the Grand Princess cruise ship off San Francisco (engaged late February 2020) which displayed the exact same largely unmutated genomic sequence (Hu-1) as determined in China during December 2019 and January 2020 [16,22].
e. The role of human-passaged (and created) regional variants lofted or plumed attached to microdust particles into the troposphere and the global wind systems, in likely attenuated form [16,28,29] see also reference in these papers to the independent assessment of the role of global wind transportation systems in past influenza pandemics in [30] Hammond et al. 1989. “Thus these authors noted the large-scale eddy circulation as causing occasional lofting and patchy deposition of virus carriers. It saw survival of the influenza virus in the air and solar radiation as important, though did not know of survival against UV in clumps, or embedded in micro-dust particles.”
All of our prior analyses and conclusions and its relation to the wider scientific literature on SARS-CoV-2/COVID-19 can be found in our past publications, which can be accessed at https://www.academia.edu/50814212/ Papers and Summary Interviews on Origin and Global Spread of COVID 19 Wickramasinghe and colleagues. The URL links to all relevant video interviews involving N. Chandra Wickramasinghe and Edward J Steele can be found in this list and at The Cosmic Tusk website of George A. Howard. https://cosmictusk.com
We have also considered and refuted the main popular explanations that were spreading uncritically abroad in both the scientific and popular media, concerning the protective efficacy of all systemic-delivered vaccines and the putative origins of COVID-19, the latter as either a jump from a latent SARS-CoV-1 animal reservoir (bat, pangolin, cat) or as a human-engineered COVID-19 genome. In the latter case this infection, identical in genomic sequence to the original Hu-1 reference (isolated in China in December 2019, NC_045512.2), was postulated to have been released from a Chinese laboratory (Wuhan Institute of Virology) either accidently or deliberately. We show both these origin explanations are scientifically implausible or impossible on genetic grounds [15,29]. Indeed, the Wuhan Lab Leak and related narratives are clearly implausible and simply do not explain what was actually observed in the first month or two of the pandemic.
Against this backdrop, we have analysed a putative “Space Weather” and “solar-wind pulse” like event [10,11] which, although poorly understood at the present time, may well account for the manner in which the pandemic signature became evident globally. At the time of writing this review the pandemic appears to have waned in severity via the natural processes of natural Herd Immunity, attenuation of the human-passaged variants and viral decay in the environment [31,32]. From the Cases per Day Curves (Figures 1-8) we think these observations have been the major new phenomenon of the pandemic that has become manifest in the data from the middle of December 2021.
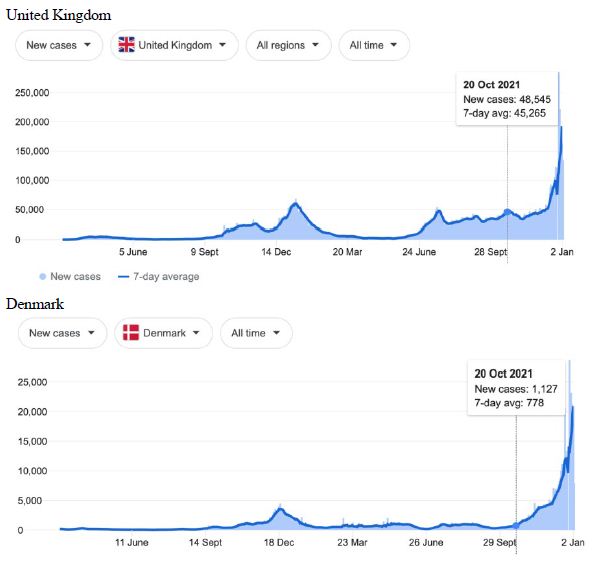
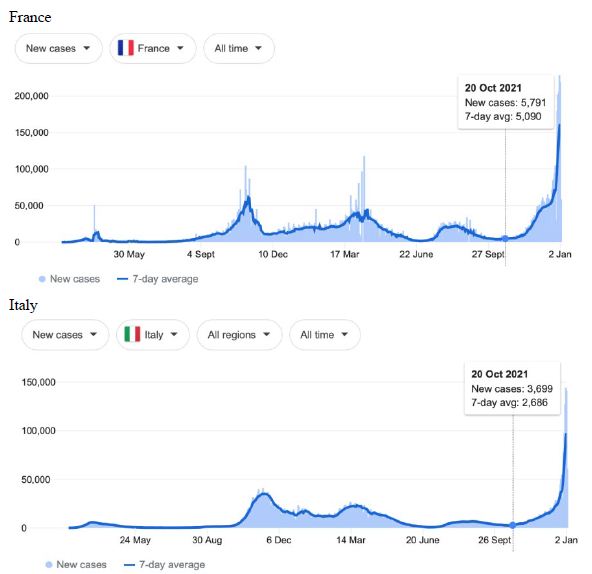
Figure 1: COVID-19 Case Rises in Selected Global Locations- Europe: United Kingdom, Denmark, France, Italy. Exponential rises in new COVID-19 cases per day as captured January 3 2022 from the Google searched site: “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser : shorturl.at/glwER
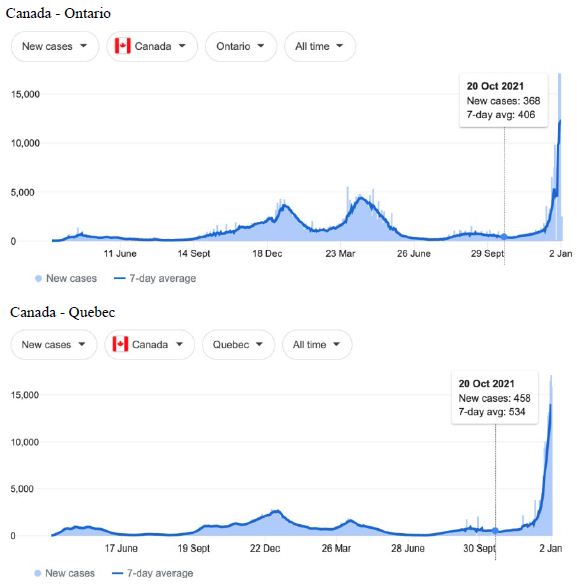
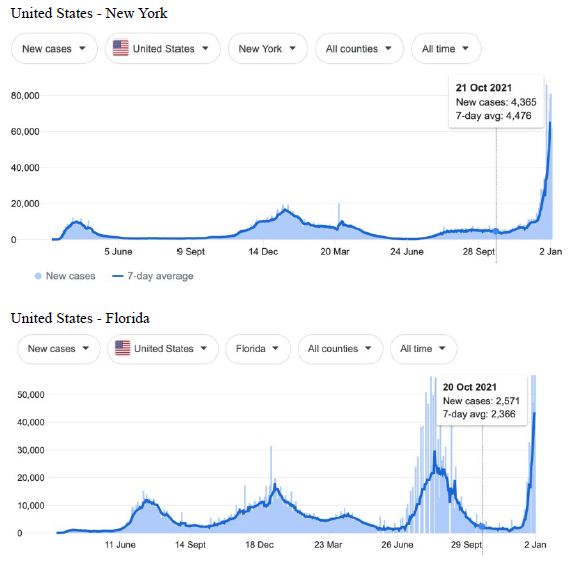
Figure 2: COVID-19 Case Rises in Selected Global Locations- Canada and USA: Ontario, Quebec, New York, Florida. Exponential rises in new COVID-19 cases per day as captured January 3 2022 from the Google searched site: “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser : shorturl.at/wQX69
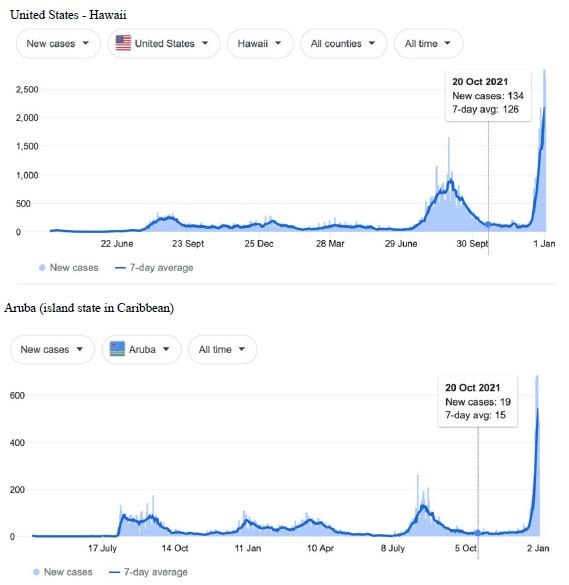
Figure 3: COVID-19 Case Rises in Selected Global Locations- Hawaii and Aruba . Exponential rises in new COVID-19 cases per day as captured January 3 2022 from the Google searched site : “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser : shorturl.at/wINR0
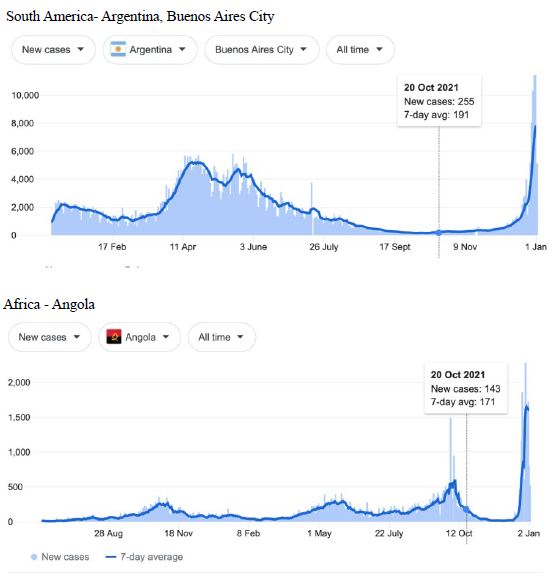
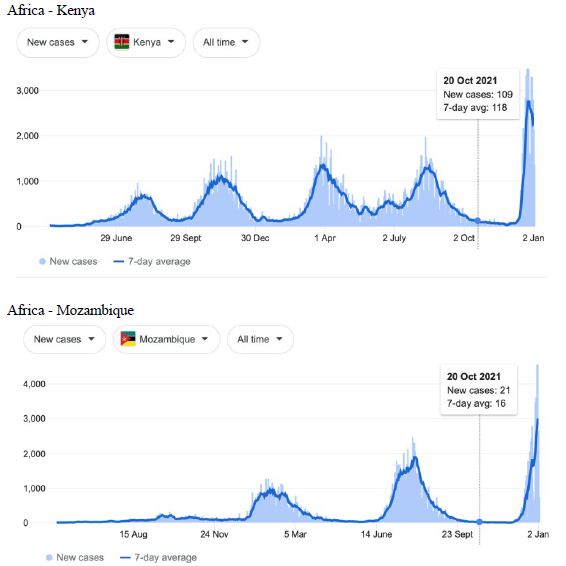
Figure 4: COVID-19 Case Rises in Selected Global Locations- South America and Africa: Buenos Aires, Angola, Kenya, Mozambique. Exponential rises in new COVID-19 cases per day as captured January 3 2022 from the Google searched site: “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser : shorturl.at/hsxGO
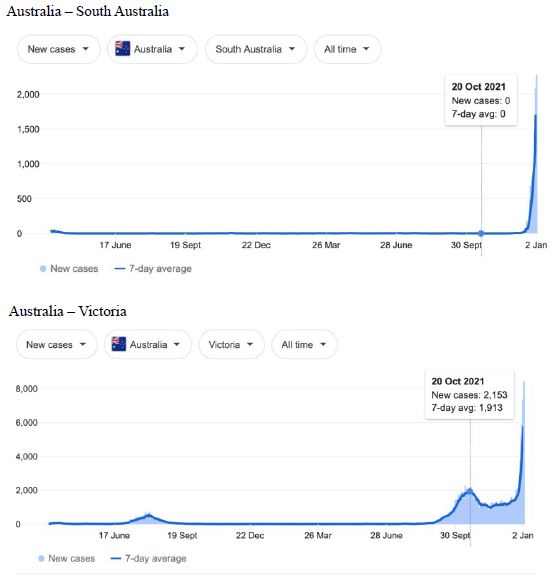
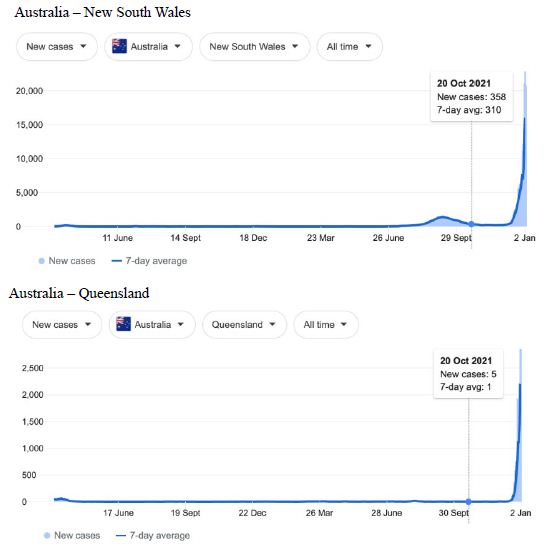
Figure 5: COVID-19 Case Rises in Selected Global Locations- Australia: South Australia, Victoria, New South Wales, Queensland . Exponential rises in new COVID-19 cases per day as captured January 3 2022 from the Google searched site: “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser : shorturl.at/lFIJ6
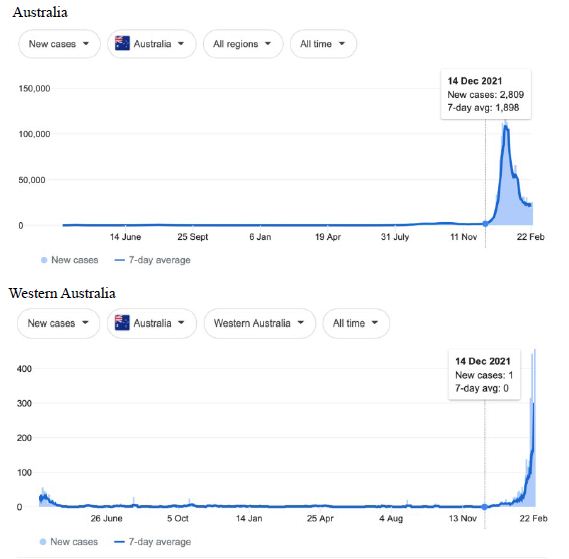
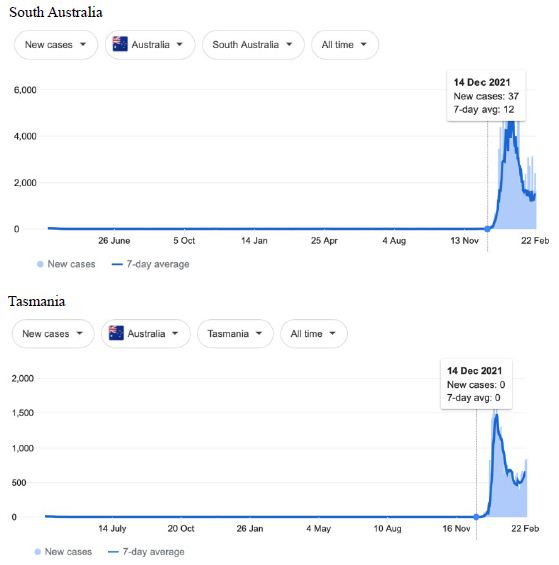
Figure 6: COVID-19 Case Rises in the whole Australia, and Western Australia, South Australia, Tasmania (similar right hand side shoulders observed for Northern Territory, Australian Capital Territory). Exponential rises in new COVID-19 cases per day as captured February 24 2022 from the Google searched site : “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser : shorturl.at/wDLTZ
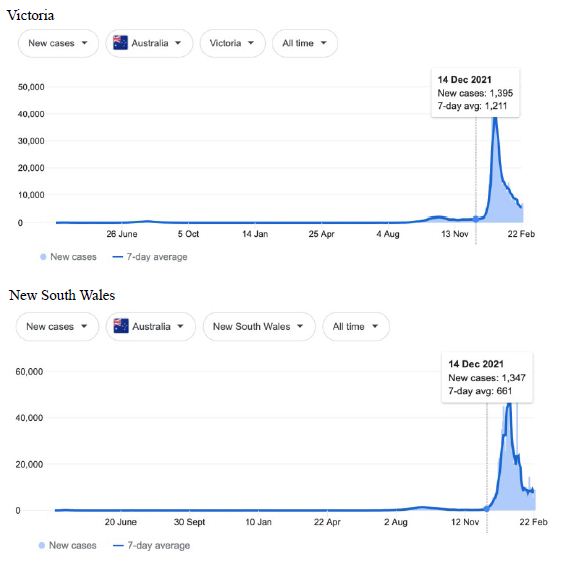
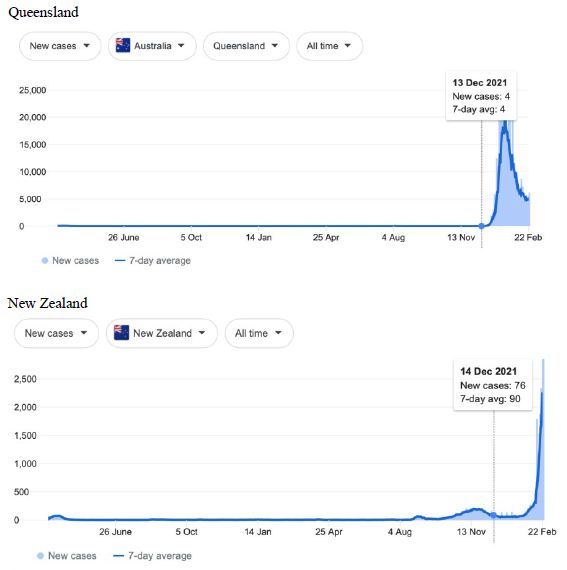
Figure 7: COVID-19 Case Rises in Victoria, New South Wales, Queensland and New Zealand (similar rising ‘hockey stick’ cases per day graph seen for French Polynesia, and Chile). Exponential rises in new COVID-19 cases per day as captured February 24 2022 from the Google searched site: “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser: shorturl.at/ersIK
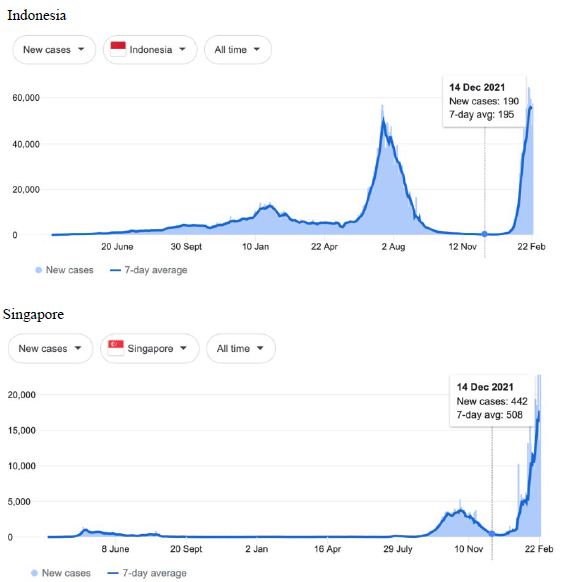
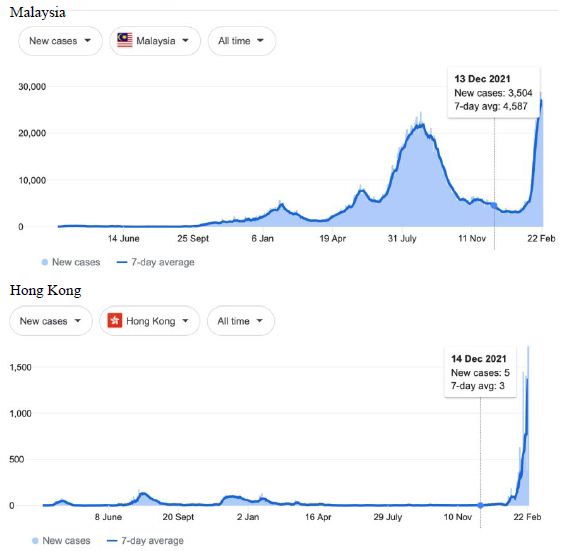
Figure 8: COVID-19 Case Rises in Indonesia, Singapore, Malaysia, Hong Kong (similar rising ‘hockey stick’ cases per day graph seen for Bhutan, Brunei, Thailand, Vietnam, Cambodia, Myanmar, South Korea, Mongolia possible shoulder in Japan). Exponential rises in new COVID-19 cases per day as captured February 24 2022 from the Google searched site: “Coronavirus disease statistics”. The URL opens at the Australia dashboard but all countries and regions can be searched via the Cases and Deaths search Menus for that region. Click or copy and paste URL into your browser : shorturl.at/lBP14
Omicron/Delta Outbreaks though December 2021 and January 2022 in Global Synchrony
Cases-per-day plots for selected locations (captured as screen shots on January 3 2022) are shown in Figures 1-5 to illustrate the extreme synchronous or simultaneous eruptions of COVID-19 epidemics (Omicron/Delta mix?) in the Northern Hemisphere regions Figures 1-3 (United Kingdom, Denmark, France, Italy, Ontario, Quebec, New York, Florida, Hawaii, Aruba), and in the Southern Hemisphere, Figures 4 and 5, embracing populated regions in South America, Africa and Australia (Buenos Aires, Angola, Kenya, Mozambique, and in Australia : South Australia, Victoria, New South Wales and Queensland). In Table 1 we list all regions of the world that display conformal exponentially rising cases per day curves over the same time interval as illustrated by the selected examples in Figures 1-5. Countries or regions with low or equivocal rises in case numbers are listed in Table 2. In some regions there was a clear peak of the presumed Omicron outbreak beginning about a week or two earlier with case numbers per day coming down in those regions (Table 3). However, many countries are ‘null zones’ with respect to this time period experiencing no rising epidemic profile (Table 4).
Table 1: Countries and Regions all showing clear Synchronous Epidemics as shown in Figures 1-5 as captured January 3 2022 (use URL Figures 1-5)

Table 2: Countries and Regions showing only a Low or Equivocal Synchronous Epidemics as shown in Figures 1-5 as captured January 3 2022 (use URL Figures 1-5)

Table 3: Countries and Regions all showing an explosive epidemic beginning a week or two earlier relative to those shown in Figures 1-5 as captured January 3 2022 (use URL Figures 1-5)
![]()
Table 4: Countries and Regions all showing no explosive epidemic or obvious begining a week or two earlier as Figures 1-5 as captured January 3 2022 (use URL Figures 1-5)
The reader can scrutinise the data at the URL site for ‘Coronavirus disease statistics’ shown in the legend to the figures. The predominant pandemic ‘strain’ evident in most regions of the world prior to these extraordinary explosive and temporally coordinated epidemic outbreaks was the ‘Delta’ strain (and related Indian-plumed strains) from the massive Indian epidemic of April-May 2021, which we hypothesized was released as a very large aerosol of many millions of trillions of virions into the troposphere for redistribution to globally distant regions via prevailing W-E, E-W and N-S wind systems [28]. The Omicron variant was found first in Botswana on November 2, 2021 and was widely assumed to have emerged first in South Africa. We discuss speculative causes of the emergence of the Omicron variant and the probable region of its origin in Section 3.
What plausible explanations can be provided for the data in Figures 1-5 and Table 1-4, and in particular the essentially simultaneous eruptions of region-specific epidemics of COVID-19 in so many different regions across the world? This is not a question that can be easily resolved. The strong indications are of a globally correlated phenomenon that we do not fully understand. One explanation could be connected to space weather events associated with the deep Sun-Spot minimum between Solar Cycles 24 and 25 [10,11]. Unseasonal weather that has been reported both in the Northern and Southern hemispheres (e.g.UK and Australia) during this time period may give a hint in this direction. The sheer numbers and global coverage of infection essentially eliminates Person-to-Person spread as the sole or main causative explanation.
A more plausible scientific explanation lies in massive region-wide in falls from the sky (the troposphere) of prior human-passaged then aerosol-plumed COVID-19 virions lofted into the troposphere and introduced into prevailing wind systems. Given current Omicron case densities, we tentatively assume a northern European origin followed by transport of viral aerosol-clouds across the Atlantic from an origin in the UK/North Europe (?), into the Pacific and Atlantic prevailing winds onto to Africa and thence to Australia.
We are still left with the conundrum of why now, and why at the same time all over the globe? The data in Figures 1-5 are but a small subset of the large number of global-wide regions in the Northern Hemisphere, Equatorial Regions, Island States, and Southern Hemisphere all struck like this at the same time (Tables 1 and 2). In addition, over this time period, many Atlantic cruise ships with double vaccinated and pre-screened passengers also became suddenly engaged with COVID-19 (assumed Omicron see [33,34]) including a fully vaccinated US Navy ship [35]. There was also the well-documented sudden outbreak from December 14 involving large numbers of fully vaccinated personnel at a remote Belgian Research Station in Antarctica thousands of miles from civilization [36]. This is indeed strikingly reminiscent of similar strikes on ships at sea and remote locations during the early phases of the pandemic in 2020 [15,16,25], including the sudden strike on the island of Sri Lanka Oct 4-5 2020 [26], and more recently on Taiwan presumed to have occurred as a result of Indian-plumed Delta virus which struck suddenly for first time from 14 May 2021 [28]. All indications are of a globally-correlated environmental trigger that we cannot fully understand at the present time.
One possible explanation is that globally dispersed viral aerosol-clouds (Omicron/Delta variant mix?) were released and lofted following human passage, and were widely distributed in the troposphere remaining viable although not immediately falling to Earth or ocean over many different regions of the world. A putative global trigger in mid-December 2021 might be postulated that brought such viral particles to earth virtually simultaneously around the world. This may have been ultimately facilitated by, but not been dependent upon, rain/precipitation [28,29]. The resulting virus-contaminated environments would then ignite outbreaks of mystery unlinked Omicron/Delta cases on a large scale giving the appearance of superfast infective spreading in a given populated contaminated region as we have previously discussed in detail for the outbreaks of mystery infections in Victoria, Australia [28,29]. This is a plausible explanation for the synchronous sudden rises of COVID-19 globally. The fact there are many “null” zones (Table 4) and ‘low’ or equivocal regions (Table 2) adds to the patchy cloud-like nature of the viral distribution in the troposphere prior to and coinciding with the solar cycle minimum. That is, it arises as deposition from the convection-driven upper troposphere which is patchy over a range of distance scales.
Another possibility, given the global nature of the present observations, and thus which cannot be ignored as a causative factor, is the known vulnerability of the Earth to pandemics during the minima of the sunspot Solar Cycle [10,11]. Thus it could be an ill-defined and poorly understood physical event broadly classed as a “Space Weather” event associated with the sunspot cycle minimum, particularly now between cycles 24 and 25, where we may be most vulnerable to “pathogen attack” from the outside the Earth: viz.
“..the Earth’s magnetosphere, and the interplanetary magnetic field in its vicinity, are modulated by the solar wind that in turn controls the flow of charged particles onto the Earth [4]. During times of sunspot minima, particularly deep sunspot minima, a general weakening of magnetic field occurs which would be accompanied by an increase in the flux of cosmic rays (GCR’s) and also of electrically charged interstellar and interplanetary dust particles”… bringing charged particles ( virus-laden dust particles) to earth. Wickramasinghe et al. 2019 [11].
Getting back to the original events in late 2019 we can advance another specific scenario on what actually happened across China which was initiated in the stratosphere over N-E China in late December 2019, and then in early January 2020 after the initial input of cometary virus-carrying dust. The virus became strongly amplified in humans across China until lock-down measures came into full force. Despite efforts to wash down the streets, the virus’s long lifetime and persistence in viable condition on dust particles enabled it to spread widely in the environment. When appropriate wind conditions arrived, the virus-carrying dust was readily swept up in tropospheric winds into the East-Asian subtropical jet stream, carrying it across the Pacific to southern USA and Western Europe. Precipitation into local wind systems thus caused infection on a state-wide and country-wide scale. In 2021 similar wind-borne viral-laden dust spread over the entire sub-continent of India causing sudden eruptions of COVID-19 infections (via PANGO variants Alpha, Kappa, and Delta at least). These country-wide sudden eruptions have been noted before although they have not been fully understood [14,16,28,29].
Finally, we should consider another putative “pulse-like” causative factor that contributed to the synchronous nature of the sudden outbreaks of Omicron/Delta infections around the world. This phenomenon was not previously explored except in the broadest of terms in our earlier publications [12,13]. It is most probably related to the fact that most viruses in their cell-to-cell infection cycle are transported as enveloped clusters of mature virions [37]. This may be important if an influence associated with “Space Weather” external to the Earth somehow triggered the liberation of smaller clusters of virions associated with tropospheric dust clouds over any given region. A dust particle of 2-3 microns could theoretically envelope 40-60 COVID-19 virions. If these were suddenly liberated to fall to ground as smaller clusters (doubletons or triplets) that would result in a ten-fold sudden increase in putative infective virion clusters floating down to contaminate a terrestrial environment. The nature of this virion liberation “trigger” is unknown (temp/pressure/radiation?) so this is still a highly speculative scenario that we present for further exploration.
However, we can now add further observations, as the manuscript was being prepared for final submission. This is again consistent with a series of “pulse-like” infection events enveloping a large fraction of the globe at the same time (prior 24-48 h). (This has been noted as January14 2022 from the “Coronavirus disease statistics” database.) The following countries appear to have been all engaged in an exponential sharp rise in COVID-19 new cases per day, a synchronous effect that was not evident in the earlier survey of January 3 2022. These countries are: Brazil, Bosnia Herzegovina, Costa Rica, Cuba, Djibouti, Egypt, French Guiana, Guatemala, Guinea-Bissau, Haiti, Hungary, Liechtenstein, Lithuania, Martinique, Maldives, Montserrat, Morocco, Monaco, North Macedonia, Norway, Pakistan, Poland, Slovenia, India, Nepal, Bhutan, Kyrgyzstan, Uzbekistan, Philippines, Japan, Romania, Sao Tome and Principe, Taiwan, Thailand, Trinidad and Tobago, Tunisia, Turks and Caicos Islands, Ukraine, Mongolia, and Venezuela. All these regions could be part of the same mid-December 2021 tropospheric in-fall process just described but show an apparent delay due to data reporting and database updating.
In addition, we surveyed plots of new cases per day from the same database as captured 24 February 2022. We detected a clear further set of global-wide synchronous eruptions of COVID-19 epidemics – presumed Omicron/Delta (mix) – in Oceania Figures 6 and 7 (all Australian states and territories, New Zealand, and French Polynesia, extending to Chile) and many countries throughout South East Asia and North East Asia (Figure 8). These synchronous eruptions we speculate have occurred by simultaneous tropospheric viral-cloud in-falls contaminating the environment of populated regions occurring most likely in early February 2022. All populated regions of Australia appear to have been struck like Western Australia (Figure 6) at the same time. WA stands out with a clear “hockey stick” infection curve as it came off a very low base. This is consistent with the evident shoulders and blips on the right hand sides of the curves for NSW, VIC, TAS, and QLD. A plausible explanation is this extended shoulder is due to this second synchronous in-fall but masked by the already high Cases per Day in these other Australian states. In a related survey of the database (captured 18-19 February 2022) we detected another set of synchronous epidemic eruptions beginning about January 10 2022 and peaking late January early February 2022, these include : Algeria, Armenia, Azerbaijan, Bangladesh, Czechia, Faroe Islands, Georgia, Iran, Iraq, Jordan, Kazakhstan, Kosovo, Latvia, Libya, Moldova, Nepal, New Caledonia, Oman, Palestine, Paraguay, Russia, Slovakia, Taiwan, Yemen.
Therefore since mid-December 2021 two further separate global-wide synchronous epidemic eruptions can be detected, from about Jan 10, then from early February 2022. We highlight in particular Western Australia in Figure 6 as that vast Australian region had never before experienced a genuine tropospheric COVID-19 epidemic strike until just recently (Figure 6) – yet that state of Australia was lock-downed repeatedly and isolated from the rest of Australia and the wider world by stringent enforced border and travel restrictions. In addition, mask mandates and social distancing regulations (crowd sizes sports events, movie theaters, churches and so on) were enforced outside homes and work places, including mandatory vaccinations for most WA workers during 2020 and 2021 despite very few if any COVID-19 cases being recorded in the state (mainly in travelers coming by road and airplane from other infected zones in Australia and from overseas who were immediately isolated and quarantined). The recent large Omicron/Delta (mix?) strike completely surprised the WA Department of Health and Medical authorities as they could not explain any of the suddenly emergent infection events recorded across the state by conventional person-to-person transmissions. The apparent ‘super spreading’ was particularly strange given the population was largely fully vaccinated.
Given these striking globally-coordinated cases per day events detected by data monitoring just COVID-19, we cannot rule out additional surprises. We speculate that other unknown dust-associated pathogens (viral, bacterial, eukaryotic spp) in the troposphere have also been brought to Earth by the same general Space Weather/Solar-pulse processes during the current Sun Spot minimum between solar cycles 24 and 25. We have laid out here a range of explanations, some over-lapping, because we are dealing with a “globally correlated phenomenon that we do not fully understand.” We have done so to ensure that as many plausible alternatives are available for further discussion and consideration in order that we may ultimately understand what happened.
Speculations How Omicron may have Arisen and Where?
All news reports in Australia, USA, South America, Africa and European countries, that are all engaged in the synchronous exponential eruptions, focus on Omicron as the main variant which is rapidly replacing Delta. From all available reports and early clinical experience, the respiratory disease severity of Omicron is less than Delta. This is consistent with death rate data (confirmed at URL links in Figures 1-5) that are consistently very low, and is approaching or remains below other estimates of death rates attributed to COVID-19 (whether Original Hu-1, Alpha, Delta and now Omicron). In approximately 0.1% of all COVID-19 exposed cases [29] death is the serious outcome mainly in the ‘Immune Defenseless Elderly Co-Morbid’ patient group. Highly vulnerable patients require administration of prompt respiratory therapies to navigate their respiratory crises that follow from the infection. Such patients often display clear deficits in innate immunity, often feeding into deficits in adaptive immunity and so are particularly at risk [38-44]. Other clinical studies [45] also show that patients in this subgroup specifically display deficits in Type I and type III interferon (IFN) inducible anti-viral immunity and thus appear ‘immune defenceless’ to coronavirus respiratory tract infections and so are at a very high risk for severe outcomes including death.
How could a human-passaged variant like Omicron arise? Omicron is clearly a derivative of “Delta” a PANGO lineage name of the L241f haplotype of the Steele-Lindley replicative-haplotype scheme (Steele and Lindley 2020) with many changes in the mRNA encoding the spike protein suggestive of mutation accrual via human passage (person to person spread, P-to-P). The analysis of how a single putative cloned variant we named “L241f.1vic” spread through aged care and nursing facilities in Melbourne, Australia beginning from about 10 May 2020 through June 2020 then erupting on scale in such facilities through July and August 2020 is very informative [23]. From the full-length genomic analysis of many thousands of publicly available SARS-Cov-2 genomes (>12,000 made available by the Victorian Dept of Health through The Peter Doherty Institute) we showed previously that there were two types of clearly identifiable patients. The first displayed unmutated versions of the virus over the entire 29903 nt genomic length. These types were particularly evident in the last two weeks of June 2020 through most of July 2020. It appeared very much like the virus was being amplified on scale in hosts that were unable to mutate the RNA virus genomes at APOBEC (cytosine to uracil) and ADAR (adenosine to inosine) deamination sites [22,23]. The second group of patients, clustering in a late August time window, displayed mutated versions of the virus, again largely at APOBEC and ADAR cytosine and adenosine deamination motifs. It was concluded thus [23] to quote conclusion directly:
The data reported herein are thus consistent with the following P-to-P infection model which is also the operational hypothesis under test: clusters of immune defenceless elderly co-morbid citizens in many aged care and nursing facilities were all systematically struck with devastating force (high infection rates and death rates), with a single unmutated (or lightly mutated) SARS-CoV-2 haplotype variant (L241f.1vic). Through late June, July, August and September in 2020, this putatively cloned variant must have been spread unimpeded by carriers who were asymptomatic or lightly symptomatic infected health- care professionals and carers working across multiple age care institutions [30-33]. The large-scale amplifications of the L241.1vic variant—instanced by the size of the multiple ‘New case’ spikes (shown in Figure 1), particularly through July 2020—could have produced many trillions of L241f.1vic virions in each location thus contaminating numerous surfaces (fomites, personal effects of all types) and could have contaminated or infected human carriers in each institution. This then fuelled the further putative quantitative dominant rapid spread of this apparently capricious L241f.1vic variant into the local community and particularly to other aged care facilities leading to further putative viral amplifications in elderly co-morbid subjects. If anything, the Victorian experience underlines why elderly co-morbid citizens require very special care, protection and therapies during cold and flu seasons [31,32].
It was then speculated that the putative highly contagious, yet clearly attenuated, “UK Mutant” (Alpha) that emerged in September in 2020 in parts of southern England was generated the same way. We also think Omicron arose by similar cycles of deaminase-mediated mutagenesis in healthy almost asymptomatic carriers, then became amplified (cloned unmutated) in Immune Defenseless Elderly Co-morbids – then after one or two more cycles via healthy intermediate ‘vectors’, infecting new cohorts of Immune Defenseless Elderly Co-Morbids where it was amplified and cloned. A plumed aerosol of literally millions on trillions of Omicron virions into the immediate troposphere and prevailing wind transportation could have easily distributed Omicron in the prevailing wind systems (e.g. from Northern Europe to South Africa where first detected). This model of alternating cycles of deaminase-mutagenesis and cloning amplification could have created a plume in a real high-density hotspot. Was it around or near UK where most Omicron have been recorded initially? At the present time we acknowledge these are speculations, but given the existence of detailed genomic records from the millions of genomes now sequenced and in computer databases, these speculations can, and will be tested in the fullness of time.
Future Vaccine Developments for Next Pandemic of Cold or Flu Respiratory Tract Pathogens?
There are many public health lessons to be learnt from the COVID-19 pandemic. Near-Earth balloon launches, stratospheric airplane and orbiting platform sampling of incoming meteorite dust have all been stressed as important early warning strategies on many previous occasions in other places (discussed recently in [9,16,29,46]. However a pandemic public health management strategy employing a more effective vaccination method needs urgent consideration. The aim should be quite different from the current very simplistic strategy of intramuscular “jab-in-the arm” vaccination (irrespective whether traditional antigens are used or the poorly safety tested mRNA expression vector vaccines). Public health vaccination which mimics natural ‘Herd’ immunity” is the desired outcome, whether to coronaviruses, influenza viruses and many other respiratory tract pathogens including bacterial ssp. that cause respiratory pneumonias and severe bronchitis.
Here we review how to optimize intranasal defective /attenuated live virus vaccination for all likely future types of pandemic respiratory viruses and finally discuss promising newly published experimental data which offers some hope for the future. We and many others have discussed the failure of the current jab in the arm intramuscular mRNA vaccines to protect against COVID-19 – yet they have been mandated by many governments and public health bureaucracies [24,29,47]. Further, the mRNA vaccines which also have high adverse effect rates are ineffective on first immunological principles because of the wrong route of delivery. The current ‘jab-in-the-arm’ route of immunization cannot possibly protect against COVID-19 infection gaining entry to and growing in mucosal cells of the respiratory tract. For that the mucosal secretory IgA antibody system needs local activation [23,24,29]. In a rush to bring COVID-19 vaccines to market, it seems that science and medicine neglected a large body of work already available on the nature of immunity and host resistance to respiratory viral infections, and how best to mimic this by vaccination, and instead became seduced by advances in 21st century molecular engineering principles into production of a vaccine whose utility and clinical efficacy even now remains unknown- more troubling is that any serious long-term sequalae are entirely unexplored. This is discussed graphically and underlined in recent interviews as well in https://youtu.be/Ijc4mjiIquk and in the Asia Pacific TV interview with Mike Ryan https://rumble.com/vmrmmq-the-origins-of-covid-19-and-why-the-vaccines-dont-work.html
Based on a wealth of scientific/immunologic data gleaned over decades, as well as experience in vaccination against respiratory viral disease, it is self-evident that protective vaccination needs to be via the oral-nasal route to activate the mucosal immune response, which is responsible for Natural Immunity and “Herd Immunity” in the population. The natural decline in the incidence of severe COVID-19 outcomes (COVID-19 associated death) was well underway before the vaccine roll out in European and USA Infection zones through 2020 and early 2021 [32]. This is brought about most effectively by effective ‘Herd Immunity’ which has been documented in a large longitudinal and population base study, conducted in Denmark through 2020 [31].
We have previously discussed the two most important forms of immunity to be activated in the mucosal cells and associated lymphoid cells of the respiratory lining. The first of these is Innate Immunity- a general elevation of these activities would strengthen the “Anti-Viral Wall” in all nasal cells and mucosal lining cells. This barrier is defective in ‘Immune Defenseless Elderly Co-Morbids’ which are the primary vulnerable group in the COVID-19 pandemic. Note that this group equates to < 1% of all infected patients see Netea et al. [41] and discussed elsewhere in detail [23] based on data from many clinical studies throughout 2020 [38-45]. In our analysis of the data >99% of the population handles COVID-19 effectively via natural immune mechanisms- to these patients it is just a “Common Cold”. Both Innate Immune Interferon Type I and III anti-viral barriers in all cells would be activated, and then adaptive acquired mucosal immunity.
Secondly, Adaptive Acquired Mucosal Immunity requires, as we discuss, the induction of dimeric secretory IgA antibodies – these antibodies are demonstrably highly avid (strong binding and thus neutralizing of toxins, viruses and adhesins preventing cell adherence and cell entry) that do not activate the Complement cascade thus do not add to “inflammatory cytokine storms.” Indeed, secretory IgA is expected to competitively block antigen binding and thus nullify the antibody-dependent enhancement (ADE) by the blood borne IgG and IgM complement fixing antibodies particularly in advanced COVID-19 infections of the elderly vulnerable group [23,24]. This sequelae of ADE pathology in vaccinated individuals who then go on to catch COVID-19 for first time is discussed more fully elsewhere [29].
To ensure that intranasal vaccination is effective it is desirable that activation of the innate immune response via the Toll receptors in addition to induction of secretory IgA against the virus. An agonist, INNA-51 of the Toll-2 receptor, was patented in 2018 (WO2018176099, Treatment of respiratory infection with a TLR2 agonist). It is currently being used in a Phase 2 trial of intranasal vaccination to prevent COVID-19 with the AstraZeneca antigen [48]. A better antigen could be an inactivated virus such as Sinovac as many more epitopes would be delivered than with AstraZeneca’s viral vector containing just the SARS-CoV-2 spike protein.
Two recent papers describing experiments in mice (a small mammal with an immune system similar to, but not identical to, humans in principle) involving intra-nasal vaccine development and assessment of efficacy in protection from disease, have now been published in December 2021. Xaio and associates [49] created a defective or harmless coronavirus that cannot replicate properly, and delivered it via the intra-nasal route so as to induce both Innate Immunity (to any other viral challenge oro-nasal) and Adaptive Immunity (that is antigen specific secretory IgA). In the other study Oh and associates [50] set up intranasal priming with influenza infection or with adjuvanted recombinant neuraminidase flu vaccine. This induced local lung-resident B cell populations that secrete protective mucosal antiviral secretory IgA. In these complimentary studies, using these different intra-nasal, mucosal lining activation strategies the workers induced both elevated pan-specific Innate Immunity as recommended by Netea and associates [41] protecting against many other unrelated respiratory track pathogens, but also the necessary dimeric secretory IgA adaptive specific immunity akin to more tradition vaccination strategies. Recent published work adds to this conclusion [51].
We would argue that studies such as these offer the hope that can now look forward to the production of easily delivered, safe and effective vaccines against many epidemic respiratory viruses, irrespective of variant or viral type, so we will be well armed in advance of the next pandemic of respiratory tract infections. This would represent a real scientific advance and a saving grace for humanity.
Acknowledgements
We thank Max Rocca, Heath Goddard, and Dayal T. Wickramasinghe for discussions and Brig Klyce also for bringing our attention to Afkhami et al 2022 [51].
Conflicts of Interest
None of the co-author team has a conflict of interest apart from understanding the scientific reasons for the origin, global spread and efficacy of human immunity to SARS-CoV-2.
Multi-author Contributions
Conceptualisation EJS, NCW, RGG, GAH: Draft writing: EJS, NCW, RMG Reading the primary clean draft: All co-authors. Active Contributions to primary draft via edits and additions: RAL, PRC, MW, SC, MKW Integration of all minor changes: EJS, NCW, RMG.
Multi-author Expertise
The areas of expertise of the multi-author team are:
Edward J. Steele: Biomedical science, Immunology, Ancestral Haplotypes, Mutagenesis, Biotechnology, Evolution, Panspermia, Cosmic Biology.
Reginald M. Gorczynski: Biomedical science, Medicine, Immunology, Evolution, Panspermia, Cosmic Biology.
Robyn A. Lindley: Biomedical science, Immunology, Biotechnology, Mutagenesis, Evolution, Panspermia, Cosmic Biology.
Patrick R. Carnegie: Biomedical science, Immunology, Biotechnology.
Herbert Rebhran: Biomedical science, Ancestral Haplotypes, Veterinary Surgeon, Cosmic Biology)
Shirwan Al-Mufti: Astronomy, Astrobiology, Astrophysics, Evolution, Panspermia, Cosmic Biology.
Daryl H. Wallis: Astronomy, Astrobiology, Scanning Electron microscopy, Meteorite Analysis, Evolution, Panspermia, Cosmic Biology.
Gensuke Tokoro: Biotechnology, Vaccines, Astroeconomics, Panspermia, Cosmic Biology.
Robert Temple: Evolution, History & Philosophy Science, Panspermia, Cosmic Biology.
Ananda Nimalasuriya: Medicine, Panspermia, Cosmic Biology.
George A Howard: Deep Earth Evolution, Archaeology, Ice Ages, Environment Restoration, Panspermia, Cosmic Biology.
Mark A. Gillman: Biomedical science, Neuropharmacology – Pain & Pleasure, Evolution, Panspermia, Cosmic Biology.
Milton Wainwright: Biomedical science, Microbiology, Balloon Loft Stratosphere Sampling, Evolution, Panspermia, Cosmic Biology.
Stephen Coulson: Astrobiology, Mathematics, Astrophysics, Panspermia, Cosmic Biology.
Predrag Slijepcevic: Biomedical science, Microbiology, Evolution, Panspermia, Cosmic Biology.
Max K. Wallis: Astrobiology, Geophysics, Space Science, Mathematics, Astrophysics, Panspermia, Cosmic Biology.
Alexander Kondakov: Astrobiology, Panspermia, Cosmic Biology.
N. Chandra Wickramasinghe: Astronomy, Astrobiology, Biomedical science, Mathematics, Astrophysics, Evolution, Panspermia, Cosmic Biology.
References
- Hoyle F, Wickramasinghe NC (1978) Life Cloud.M. Dent Ltd, London.
- Hoyle F, Wickramasinghe NC (1979) Diseases from Space. J.M. Dent Ltd, London.
- Hoyle F, Wickramasinghe NC (1981) Evolution from Space. J.M. Dent Ltd, London.
- Hoyle F, Wickramasinghe NC (1985) Living Comets. Univ. College, Cardiff Press, Cardiff.
- Hoyle F, Wickramasinghe NC (1991) The Theory of Cosmic Grains. Kluwer, Dordrecht.
- Hoyle F, Wickramasinghe NC (1993) Our Place in the Cosmos: the Unfinished Revolution. J.M. Dent Ltd, London.
- Hoyle F, Wickramasinghe NC (1999) Astronomical Origins of Life: Steps towards Panspermia. Reprints from Astrophys. Space Sci 268: 1-3. Kluwer Academic Publishers, Dordrecht, The Netherlands.
- Wickramasinghe C (2020) Diseases from Outer Space: Our Cosmic Destiny. 2nd Edition of Diseases from Space, World Scientific Publishing Company, Singapore.
- Wainwright M, Rose CE, Baker AJ, Wickramasinghe NC, Omairi T (2015) Biological Entities Isolated from Two Stratosphere Launches-Continued Evidence for a Space Origin. Astrobiol Outreach 3:2. https://www.walshmedicalmedia.com/archive/jao-volume-3-issue-2-year-2015.html
- Wickramasinghe NC, Steele EJ, Wainwright M, Tokoro G, Fernando M, et al. (2017) Sunspot Cycle Minima and Pandemics: The Case for Vigilance? J Astrobiol Outreach 5:2. https://www.walshmedicalmedia.com/open-access/sunspot-cycle-minima-and-pandemics-the-case-for-vigilance-2332-2519-1000159.pdf
- Wickramasinghe NC, Wickramsainghe DT, Senanayake S, Qu J, Tokoro G, et al. (2019) Space Weather and Pandemic Warnings? Sci 117: 1554. http://www.lifefromspace.com/resources/CurrentScience2020.pdf
- Steele EJ, Al-Mufti S, Augustyn KA, Chandrajith R, Coghlan JP, et al. (2018) Causes of Cambrian Explosion-Terrestrial or Cosmic? Biophys Mol Biol 136: 3-23. [crossref]
- Steele EJ, Gorczyski RM, Lindley RA, Liu Y, Temple R, et al. (2019 ) Lamarck and Panspermia-On the efficient spread of living systems throughout the cosmos. Prog Biophys. Mol. Biol 149: 10-32. [crossref]
- Steele EJ, Gorczynski RM, Lindley RA, Tokoro G, Temple R, et al. (2020) Origin of new emergent Coronavirus and Candida fungal diseases- Terrestrial or Cosmic? Genetics 106: 75-100. [crossref]
- Steele EJ, Gorczynski RM, Rebhan H, Carnegie P, Temple R, et al. (2020) Implications of haplotype switching for the origin and global spread of COVID-19. Virology: Current Research 4: 2020. DOI: 37421/Virol Curr Res.2020.4.115
- Steele EJ, Gorczynski RM, Lindley RA, Tokoro G, Wallis DH, et al. (2021) Cometary Origin of COVID-19 Infect Dis Ther 2: 1-4. https://researchopenworld.com/ cometary-origin-of-covid-19/ DOI: 31038/IDT.2021212
- Steele EJ, Wickramasinghe NC (2020) (Eds). Cosmic Genetic Evolution Academic Press-Elsevier: Advances in Genetics. Volume 106, Serial Editor Dhavendra Kumar, London and San Diego. https://www.elsevier.com/books/cosmic-genetic-evolution/steele/978-0-12-821518-0
- Wickramasinghe NC, Steele EJ, Gorczynski RM, Temple R, Tokoro G, et al. (2020) Comments on the Origin and Spread of the 2019 Coronavirus. Virology: Current Research 4: 1. DOI: 37421/vcrh.2020.4.109
- Coulson SG, Wickramasinghe NC. (2003) Frictional and radiation heating of micron-sized meteoroids in the Earth’s upper atmosphere. Not. Roy. Astron. Soc 343: 1123-1130.
- Wickramasinghe NC, Steele EJ, Gorczynski RM, Temple R, Tokoro G, et al. (2020) Growing Evidence against Global Infection-Driven b Person-to-Person Transfer of COVID-19. Virology Current Research 4:1. DOI: 37421/vcrh.2020.4.110
- Wickramasinghe NC, Steele EJ, Gorczynski RM, Temple R, Tokoro G, et al. (2020) Predicting the Future Trajectory of COVID. Virology Current Research 4:1. DOI: 37421/vcrh.2020.4.111
- Steele EJ, Lindley RA (2020) Analysis of APOBEC and ADAR deaminase-driven Riboswitch Haplotypes in COVID-19 RNA strain variants and the implications for vaccine design. Research Reports https://www.companyofscientists.com/index.php/rr
- Lindley RA, Steele EJ (2021) Analysis of SARS-CoV-2 haplotypes and genomic sequences during 2020 in Victoria, Australia, in the context of putative deficits in innate immune deaminase anti-viral responses. Scand J Immunol. https://doi.org/10.1111/sji.13100
- Gorczynski RM, Lindley RA, Steele EJ, Wickramasinghe NC (2021) Nature of Acquired Immune Responses, Epitope Specificity and Resultant Protection from SARS-CoV-2. Pers. Med 11: 1253. https://doi.org/10.3390/jpm11121253
- Howard GA, Wickramasinghe NC, Rebhan H, Steele EJ, Gorczynski RM, et al (2020) Mid-Ocean Outbreaks of COVID-19 with Tell-Tale Signs of Aerial Incidence. Virology Current Research 4:1. DOI: 37421/vcrh.2020.4.114
- Wickramasinghe NC, Steele EJ, Nimalasuriya A, Gorczynski RM, Tokoro G, et al. (2020) Seasonality of Respiratory Viruses Including SARS-CoV-2. Virology Current Research 4:2. DOI: 37421/vcrh.2020.4.117
- Wickramasinghe NC, Wallis MK, Coulson SG, Kondakov A, Steele EJ, et al. (2020) Intercontinental Spread of COVID-19 on Global Wind Systems. Virology Current Research 4:1. DOI: 37421/vcrh.2020.4.113
- Steele EJ, Gorczynski RM, Carnegie P, Tokoro G, Wallis DH, et al (2021) COVID-19 Sudden Outbreak of Mystery Case Transmissions in Victoria, Australia, May-June 2021: Strong Evidence of Tropospheric Transport of Human Passaged Infective Virions from the Indian Epidemic. Infect Dis Ther 2: 1-28. DOI: 31038/IDT.2021214
- Steele EJ, Gorczynski RM, Rebhan H, Tokoro G, Wallis DH, et al. (2021) Exploding Five COVID-19 Myths On the Origin, Global Spread and Immunity. Infect Dis Ther 2: 1-15. DOI: https://doi.org/10.31038/IDT.2021223
- Hammond GW, Raddatz RL, Gelskey DE (1989) Impact of atmospheric dispersion and transport of viral aerosols on the epidemiology of Influenza. Reviews of Infectious Disease 11: 494-497. [crossref]
- Hansen CH, Michlmayr D, Gubbels SM, Mølbak K, Ethelberg S (2021) Assessment of protection against reinfection with SARS-CoV-2 among 4 million PCR-tested individuals in Denmark in 2020: A population-level observational study. The Lancet 397: 1204-1212. DOI: https://doi.org/10.1016/S0140-6736(21)00575-4
- Steele EJ, Gorczynski RM, Lindley RA, Tokoro G, Wallis DH, et al. (2021) An End of the COVID-19 Pandemic in Sight? Infect Dis Ther 2: 1-5. DOI: 31038/IDT.2021222
- Lee H. (2021) CDC Investigates 86 Cruise Ships With COVID-19 Outbreaks Dec 28 2021. https://www.theepochtimes.com/cdc-investigates-86-cruise-ships-with-covid-19-outbreaks_4181845.html
- Khalip A, Pereira M (2021) COVID outbreak ends cruise for thousands on German ship in Lisbon https://www.reuters.com/world/europe/covid-outbreak-ends-cruise-thousands-german-ship-lisbon-2022-01-02/
- AP|Washington 2021 (Dec 25) US Navy pauses warship deployment to South America amid Covid-19 outbreak. https://www.business-standard.com/article/international/us-navy-pauses-warship-deployment-to-south-america-amid-covid-19-outbreak-121122500109_1.html
- BBC News Coronavirus Pandemic: Antarctic Outpost hit by Covid-19 outbreak. https://www.bbc.com/news/world-europe-59848160
- Combe M, Garijo R, Geller R, Cuevas JM, Sanjaun R (2015) Single-cell analysis of RNA virus infection identifies multiple genetically diverse viral genomes within single infectious units. Cell Host Microbe 18: 424-432. [crossref]
- Achary D, Liu G-Q, Gack MU (2020) Dysregulation of type I interferon responses in COVID-19. Rev Immunol 20: 397- 398. [crossref]
- Blanco-Melo D, Nilsson-Payant BE, Liu W-C, Uhl S, Hoagland D, et al. (2020) Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 181: 1036-1045. [crossref]
- Hadjadj J, Yatim N, Barnabei L, Corneau A, Boussier J, et al. (2020) Impaired type I interferon activity and exacerbated inflammatory responses in severe Covid-19 patients. Science 369: 718-724. DOI: 1126/science.abc6027
- Netea MG, Giamarellos-Bourboulis EJ, Domınguez-Andre’s J, Curtis N, Reinoutvan C, et al. (2020) Trained Immunity: A tool for reducing susceptibility to and the severity of SARS-CoV-2 infection. Cell 181: 969-977. [crossref]
- Zhang Q, Bastard P, Liu Z, Le Pen J, Moncada-Velez M, et al. (2020) Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 370: eabd4570. [crossref]
- Moderbacher CR, Ramirez SI, Dan JM, Grifron A, Hastie KM, et al. (2020) Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell 183: 996-1012. [crossref]
- Sette A, Crotty S (2021) Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 184: 861-880. [crossref]
- Lucas C, Wong P, Klein J, Castro TBR, Silva J, et al. (2020) Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 584: 463-469. [crossref]
- Qu J, Wickramasinghe NC (2020) The world should establish an early warning system for new viral infectious diseases by space-weather monitoring. MedComm 1: 423 -426. [crossref]
- Subramanian SV, Kumar A. (2021) Increases in COVID-19 are unrelated to levels of vaccination across 68 countries and 2947 counties in the United States. Eur J Epidemiol 36: 1237-1240. https://doi.org/10.1007/s10654-021-00808-7
- Deliyannis G, Wong CY, McQuilten HA, Bachem A, Clarke M, et al. (2021) TLR2-mediated activation of innate responses in the upper airways confers antiviral protection of the lungs. JCI Insight 6: e140267. [crossref]
- Xiao Y, Lidsky PV, Shirogane Y, Aviner R, Wu CT, et al. (2021) A defective viral genome strategy elicits broad protective immunity against respiratory viruses. Cell 184: 6037-6051. [crossref]
- Oh JE, Song E, Moriyama M, Wong P, Zhang S, et al (2021) Intranasal priming induces local lung-resident B cell populations that secrete protective mucosal antiviral IgA. Science Immunology 6: eabj5129. [crossref]
- Afkhami S, D’Agostino MR, Zhang A, Stacey HD, Marzok A, et al. (2022) Respiratory mucosal delivery of next-generation COVID-19 vaccine provides robust protection against both ancestral and variant strains of SARS-CoV-2. Cell 185: 896–915. [crossref]