Abstract
IODP site U1376 was drilled on Burton Guyot on the Louisville Seamount Trend in the South Pacific. Site U1376 encountered an algal reef limestone from 23.45-38.60 mbsf immediately overlying a 3 m thick basalt pebble conglomerate above the volcanic basement rocks in the area. This limestone section was reported as Cretaceous in age by the shipboard party based on possible late Cretaceous rudist fossils in the underlying conglomerate, but planktonic foraminifera suggest a Paleogene (possibly Eocene) age. Paleogene algal reefs are of interest as many have rhodophyte and macroforaminifera as the framework builders of the reef (foralgal reefs) and many represent an equilibrium reef stage during global warmth. Most known foralgal reefs such as the Salt Mountain Limestone of Alabama developed along continental shelves. Others such as Red Gal Ring in Jamaica and the Uitoé Limestone in New Caledonia developed within active tectonic zones. Site U1376 represents a unique opportunity to examine a foralgal reef which developed in isolation. The limestone section in site U1376 (referred to henceforth informally as the Burton Guyot limestone) consists of 15.15 m of rhodophyte coralline algal boundstones. Assemblages of both crustose and frondose forms of the rhodophytes occur in five distinct zonations within the Burton Guyot limestone. The zones of frondose rhodophytes represent a response to increasing accommodation (catch-up growth) produced by some combination of subsidence and rising eustatic sea level. The zones of crustose rhodophytes in the core represent the growth response during high stand (keep-up growth) and likely represent limited available accommodation space. Erosional surfaces at the tops of the crustose zones represent sea level change. In the initial shipboard description of the Burton Guyot limestone, only isolated foraminifera was reported. Examination of the thin-sections revealed numerous foraminifera. Planktonic foraminifera are more abundant than benthonic, and macroforaminifera are less abundant than in other foralgal reefs. In contrast to the foralgal reefs in the Gulf Coastal Plain, Jamaica, and New Caledonia, macroforaminifera play only a minor role in the framework of the reef structure. The planktonic foraminifera identified are Subbotina eocaena, Catapsydrax univcaus, Catapsydrax sp., Globorataloides quadrocarmeratus, Parasubbotina eocclova, Globigerina officinalis, Parasubbotina varianata, Turborotalia pomeroli, and Turborotalia frontosa. This assemblage indicates an age of middle Eocene (late Lutetian 41 Ma).
The middle Eocene foralgal reef that developed on Burton Guyot has a much simpler sedimentary architecture than the other foralgal reefs studied. This may be due to a dominant role of subsidence in the creation of accommodation on Burton Seamount. It may also be due to the direct interaction of oceanic process with the reef framework builders as opposed to interaction with continental margin processes. Further study of the Burton Guyot limestone may refine the paleoecologic controls on the development of isolated reefs during the Paleogene.
Introduction
The Integrated Ocean Drilling Program (IOPD) site U1376 was drilled on Burton Guyot as a part of Expedition 330 on the Louisville Sea Mount Trail in the Southwest Pacific (Figure 1). During drilling, an algal reef limestone (Subunit IIA of the expedition 330 Scientist, 2012) was encountered immediately overlying the volcanic basement units and a thin (3.3 m) basalt conglomerate. The algal reef limestone unit (here in referred to informally as the Burton Guyot limestone) is 15.15 meters in thickness and is overlain by shallow water volcanic sandstone and breccias. The unit is underlain by 3.3 m thick heterolithic basalt conglomerate lying directly on the volcanic basement. The Burton Guyot limestone is primarily a rhodophyte boundstone. The age of the Burton Guyot limestone is uncertain, having been reported as late Cretaceous by the Expedition 330 Scientist (2012) [1]. The age assessment was inferred by the presence of rudist fragments in the underlying conglomerate bed. The lack of rudists in the Burton Guyot limestone suggests a younger (Paleogene) in age. Koppers et al. (2012) suggested an age of ~65 Ma for the volcanic basement of the Burton Seamount. This is a Paleocene (Danian) age according to Gradstein et al. 2012 [2] and 2020 [3] and indicates that the Burton Guyot limestone is younger. Preliminary macroscopic examination of samples of the Burton Guyot limestone indicated the presence of both crustose and frondose rhodophytes. Additionally, some macroforaminifera were seen in the samples. The presence of these fossils and the inferred Paleogene age suggests the possibility that the Burton Guyot limestone may have originated as a foralagl reef. A foralgal reef is a bioherm where rhodophytes and macroforaminifea are the primary framework builders. They are the dominant type of reef found in the Paleocene and Eocene rocks worldwide. Foralgal reefs are of interest as they may represent a reef type which is in equilibrium with greenhouse climatic condition. Foralgal reefs may represent a model for future change in Neogene and Quaternary coral reefs. Some of the better known foralgal reefs include the Paleocene Salt Mountain Limestone of Alabama (Toulmin, 1941; Bryan et al. 1997) [4], the middle Eocene Red Gal Ring section in Jamaica (Robinson, 1974) and the middle Eocene Uitoe Limestone of New Caledonia (Harrison, 2013). While these three foralgal reefs are known to respond to change in relative sea level change, all three are in settings where the preserved eustatic signal is weak. The Salt Mountain foralgal reef developed at lowstand in a coastal plain setting influenced by salt tectonics. Both the Red Gal Ring and Uitoe foralgal reef s developed in active convergent tectonic settings. By contrast, the foralgal reef of the Burton Guyot limestone developed in isolation with accommodation produced by subsidence associated with Louisville Seamount Trail hotspot. Sequential change in the biofacies identified within the Burton Seamount limestone should record changes sea level associated with subsidence of the Louisville Seamount Trail and eustasy.
It is the purpose of this paper to examine thin sections of the Burton Guyot limestone for foraminifera with the goals of:
- Correlating this limestone to the geologic time scale of Gradstein et al. (2012) [2] via planktonic foraminiferal
- Describe the succession of the biofacies present in the Burton Guyot limestone to produce a relative sea level curve for the
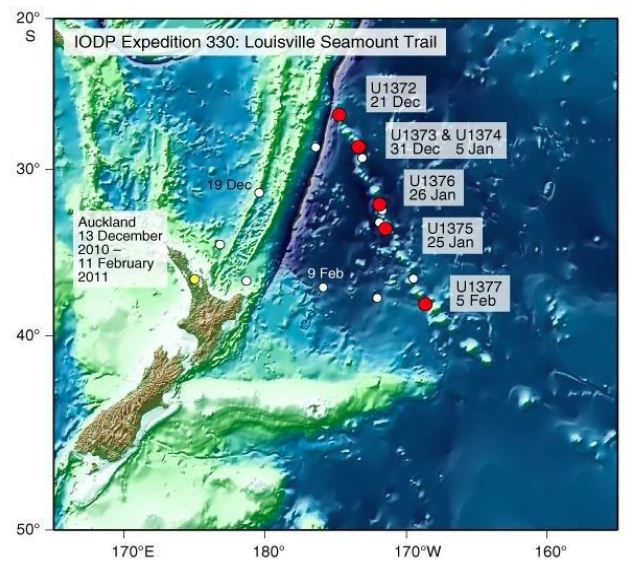
Figure 1: The general location of the study area [5]
Geologic Setting
The Louisville Seamount Trail is a Southeast-Northwest linear trend of seamounts located on the southwestern Pacific Plate from approximately 45° 32’ S, 157° 23’ W to the convergent boundary with the Australian Plate. OPD Expedition 330 the trend to better understand better the relationship between the geologic history of the Louisville Seamount Trail and the Hawaiian –Emperor Seamount Trail and whether a motion on two associated hotspots moved in concert (IOPD Expedition 330, 2012) [1]. IOPD site U1376 on Burton Guyot was the only site among the six drilled in Expedition 330 which encountered biohermal limestone. The Burton Guyot limestone is found from 23.45 meters below seafloor (mbsf) to 38.60 mbsf in site U1376 (Figure 2). The lithologic characteristics and core recovery of this unit did not permit an even sampling interval. A total of 22 samples were collected, and thin sections were prepared. These thin sections are housed in the biostratigraphy Laboratory at Ball State University. The sample location in the core and the lithologic description of each thin section are shown in Table 1. The Burton Guyot limestone was identified by Expedition 330 Scientists (2012) [1,5] as rhodophyte boundstone. While rhodophytes are abundant, detailed examination of the Burton Guyot limestone in thin sections reveals a diverse suite of carbonate lithologies. Wackestone, packstone, and grainstone are present in the thin section. All these lithologies contain abundant fossils fragments. Both crustose and frondose forms of the rhodophytes occur throughout the Burton Guyot limestone. In addition to the rhodophytes, abundant by fossil fragments of echinoderms, pelecypods, gastropods, ostracodes, and coral are present. Planktonic foraminifera, small benthic foraminifera, and macroforaminifera are rare throughout the core with the planktonic foraminifera being the most abundant. The detailed lithology of the studied interval is shown in Figure 1.

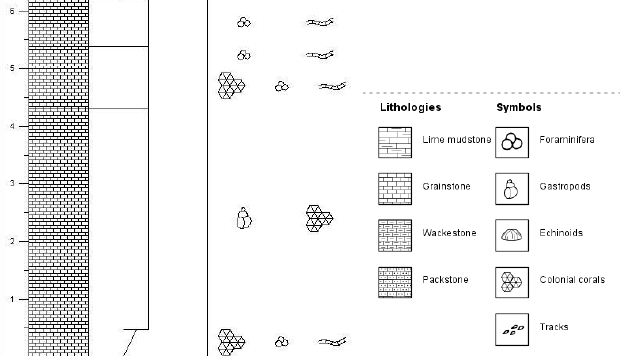
Figure 2: The columnar section of the study area
Table 1: Description of the core samples
|
Location/Location IOPD Expedition 330 -site U1376 |
Code Side |
Sample NO |
Depth |
Dunham Classification |
Lithology Description |
| 3R 4W
31/33 |
1 |
0.69 cm |
Mudstone | Yellow to a creamy color, medium hard. | |
| 3R 4W
108/110 |
2 |
0.32 cm |
Grainstone | White color, hard, coral fragments, and fossils fragments. | |
| 3R 4W
133/135 |
3 |
1 m |
Grainstone | White to yellow color, hard, coral fragments.
Subbotina eocaena |
|
| 3R 5W
40/42 |
4 |
0.28 cm |
Grainstone | White color, hard, Coral sp?, fossils fragments.
Alabamina sp., |
|
| 3R 5W
98/100 |
5 |
0.3 cm |
Grainstone | White color, hard, fossils fragments. | |
| 3R 5W
135/137 |
6 |
1 m.166 cm |
Pack-Grainstone | White color, hard, and fossils fragments.
Catapsydrax univcaus. Catapsydrax sp., |
|
| 3R 6W
37/38 |
7 |
0.68 cm |
Grainstone | White color, hard, fossils fragments.
Heterostegina sp., |
|
| 3R 4W 4/6 |
8 |
2 m.204 cm |
Pack-Grainstone | Yellow to brown color, medium hard | |
| 4R 1W
25/27 |
9 |
0.69 cm |
Pack-Grainstone | White color, hard, fossils fragments.
Catapsydrax univcaus. |
|
| 3R 6W
4/5 |
10 |
0.75 cm |
Grainstone | White color, medium hard, fossiliferous. Note: black grains might be resulted from leaching igneous rock | |
| 4R 2W
20/22 |
11 |
0.26 cm |
Boundstone | White to brown color, hard, a trace of fossil fragments.
Parasubbotina varianata |
|
| 4R 2W
45/47 |
12 |
1 m.08 cm |
Pack- Grainstone | White to brown color, medium hard, traces of gastropod. | |
| 4R 3W
6/8 |
13 |
1 m |
Grainstone | White color, medium hard, fossiliferous.
Globorataloides quadrocarmertus Parasubbotina eocclova Lagena sp., Discocyclina ( Discocyclina) marginata? |
|
| 4R 3W
96/98 |
14 |
0.41 cm |
Wack- Packstone | White color, hard, fossils fragments.
Parasubbtina ecoclava. |
|
| 4R 4W
5/6 |
15 |
0.50 cm |
Grainstone | White to brown color, medium hard Turborotalia pomeroli Cibicidoides micrus | |
| 4R 4W
27/29 |
16 |
0.78 cm |
boundstone | White color, medium hard, fossiliferous.
Globigerina officinaliss |
|
| 4R 4W
90/92 |
17 |
0.35 cm |
Grainstone | White color with pink spots, medium hard, fossil fragments | |
| 4R 4W
75/77 |
18 |
0.5 cm |
Grainstone | Yellow to brown Color, medium hard, calcite grains appearance.
Paragloborotalia (Turborotalia)griffinoides |
|
| 5R 1W
5 |
19 |
0.97 cm |
Mud-Wackstone | Gray to black color, very hard. | |
| 5R 1W
8/10 |
20 |
0.64 cm |
Packstone | White color, medium hard, abundant by gastropods and fossils fragments. | |
| 5R 1W
77/79 |
21 |
0.97 cm |
Wack-Pacstone | Brown to Black color, very hard.
Turborotalia frontosa |
|
| 5R 1W
95/96 |
22 |
1m |
Mudstone to Wackstone | Gray to black color, medium hard |
Results and Discussions
Abundance and Stratigraphic Distribution of Foraminifera, Site-U1376
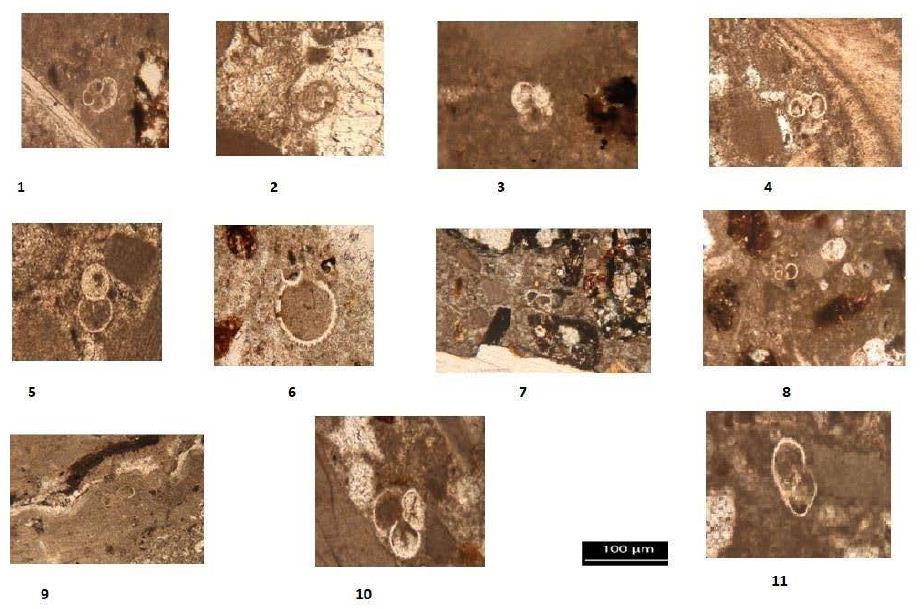
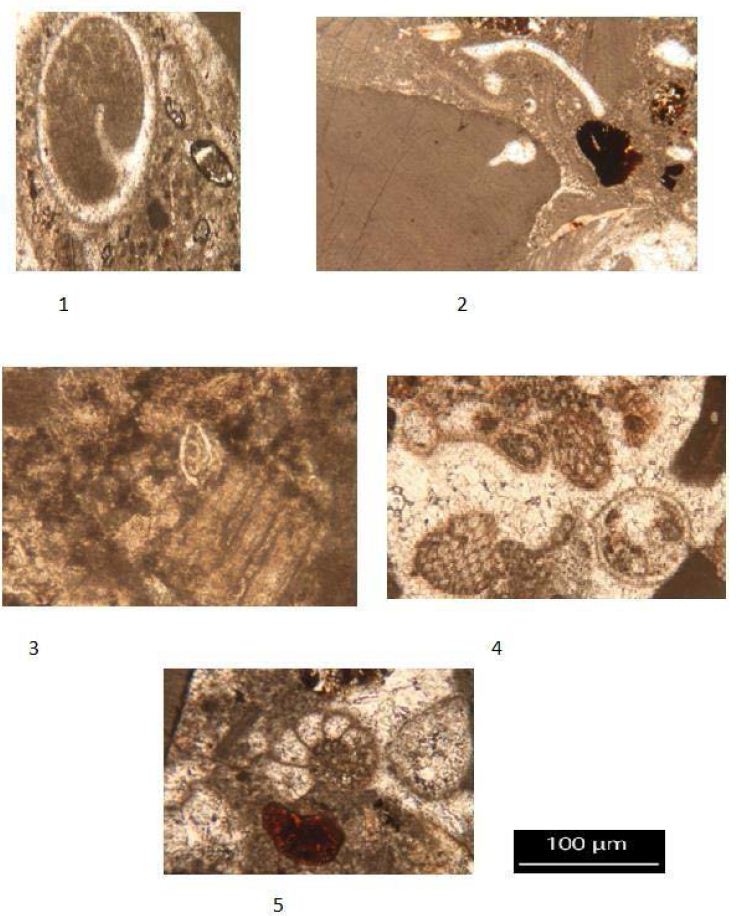
Planktonic foraminifera were identified in thin-section throughout the Burton Guyot limestone. Ten species were identified in the studied interval. They include Catapsydrax unicavus, Catapsydrax sp., Globorotaloides quadrocameratus, Paragloborotalia griffiniodes, Parasubbbotina eoclava, Parasubbbotina varianta, Globigerina officinalis, Subbbotina eocaena, Turborotalia frontosa and Turborotalia pomeroli. Planktonic foraminifera are rare in the Burton Guyot limestone occurring in only 10 of the 22 studied thin-sections. Additionally, most of the species are represented by only a single specimen. The distribution of the planktonic foraminifera from the Burton Guyot limestone is shown in Table 2. Some benthic foraminifera were found in the thin-sections. These have been identified as Alabamina sp., Cibicidoides micrus, and Lagena sp., all long ranging taxa. Two specimens of macroforaminifera (Discocyclina marginata and Heterostegina sp,.) were also found in the thin-sections of the Burton Guyot limestone. The low number of macroforaminifera was surprising as other foralgal reefs around the world contain macroforaminifera as a conspicuous part of the fauna. The distribution of smaller benthic foraminifera and macroforaminifera in the Burton Guyot limestone are shown in Table 3. Assemblages of both crustose and frondose forms of the rhodophytes occur in five distinct zonations within the Burton Guyot limestone. The zones of frondose rhodophytes represent a response to increasing accommodation space (catch-up growth) produced by some combination of subsidence and rising eustatic sea level as shown in Figure 2.
Foraminiferal assemblages, it is only a minor component of those assemblages. Isotopic studies of C. unicavus indicate it occupied a deep planktonic habitat and its relative abundance at Burton Guyot are consistent with an environment characterized by subsidence. By comparison, the foraminifera from the other Paleogene foralgal reefs in the Salt Mountain Limestone of Alabama and the Uitoé Limestone of New Caledonia, are relatively more abundant in their respective sections as shown in Figure 3.
Table 2: Distribution of biostratigraphically important foraminifera in the Burton Guyot limestone
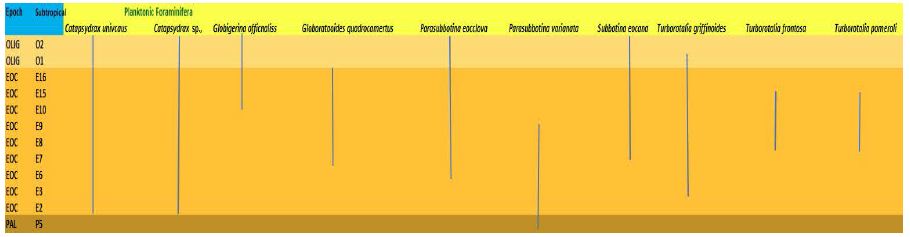
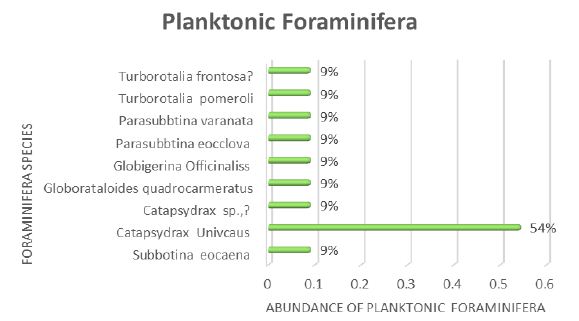
Figure 3: The abundance and stratigraphic distribution of planktonic foraminifera at Site – U1376, in the form of a frequency diagram. The y-axis represents the species name, and the x-axis represents the percentage of species.
Across the LLTM (The Late Lutetian Thermal Maximum) Benthic foraminifera species were not extinct, but mild assemblage shifts indicate environmental disturbances, perhaps due to variations in the type of organic materials transported to the seafloor. Based on thin sections from this core, the representative benthic foraminifera assemblages are less prevalent in this section, with a distribution of less than 1% This provides insight into how the depth of the sea level has changed and been impacted by it as seen in Figure 4.
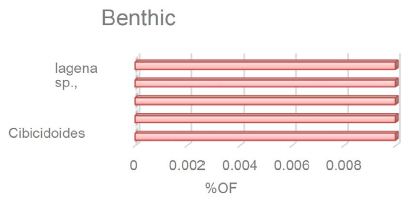
Figure 4: The stratigraphic distribution and abundance of benthic foraminifera at Site U1376. Note that the x-axis displays the proportion of species.
Stratigraphic Range of Planktonic and Benthic Foraminifera (Eocene) Species
The two next figures show the age estimate of both planktonic and benthic foraminifera based on the biohrizonsLourens et al. 2004, which confirmed all the species are Eocene in age, as shown in Figures 5 and 6.

Figure 5: Stratigraphic range of planktonic (Eocene) species
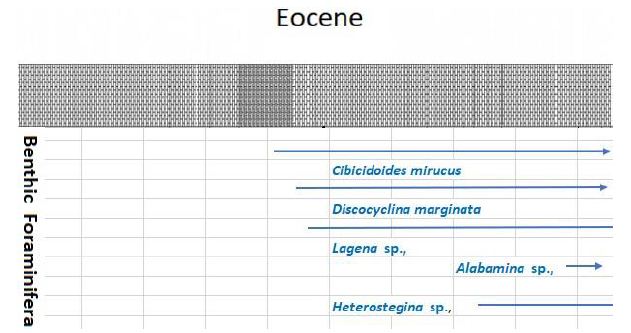
Figure 6: Stratigraphic range of benthic foraminifera (Eocene) species
Biostratigraphy
The biozonation used in this study is that of Berggren [6] with modifications of Wade et al. (2011). he greatest number of samples containing planktonic foraminifera was obtained from the Burton Guyot Limestone Unit. At this locality, only nine biozones from the Paleocene and lower Eocene were recognized with certainty. The age of this unit based on the overlapping ranging of the rare planktonic foraminifera is Middle Eocene (late Lutetian- 47.8 Ma) in age as shown in Table 2.
Taxonomy of Foraminifera Planktonic Foraminifera
Order Foraminiferida (Eichwald, 1930)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Globigerinidae(Carpenter, Parker and Jones, 1862)
1- Subbotina eocaena (Guembel, 1968)
These species have a large size subbotiondes during the Eocene.
Stratigraphic range Zone E6?- to Zone O1. Early Eocene to Early Oligocene [6]
Geographic distribution at the middle latitude range.
Order Foraminiferida (Eichwald, 1930)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Globigerinidae(Carpenter, Parker and Jones, 1862)
2- Catapsydrax Univcaus (Bolli, Loblich, and Tappan, 1957) [7]
These species have a large size represent (only) in the Middle to Upper Eocene.
Stratigraphical range Zone E2 to Zone N6.
Geographical distribution at the Global range.
Order Foraminiferida (Eichwald, 1930)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Globigerinidae(Carpenter, Parker and Jones, 1862)
3- Globorataloides quadrocameratus [6]
Stratigraphic range Zone E2 to E16.
Geographic distribution these species have widely distributed in the tropical and high latitude.
The Age is Early Eocene to Early Oligocene.
Order Foraminiferida (Eichwald, 1930)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Globigerinidae(Carpenter, Parker and Jones, 1862)
4- Parasubbotina eocclova (Coxall et al. 2003)
Age Middle Eocene [6]
Stratigraphic Range Zone E7 to E9 (Coxall et al. 2003).
Geographic distribution; Theses species have widely distributed in the low to Middle latitudes.
Order Foraminiferida (Eichwald, 1930)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Globigerinidae(Carpenter, Parker and Jones, 1862)
5- Globigerina Officinaliss [8]
Age Middle Eocene to Oligocene [6]
Stratigraphical Range Zone E10 to Oligocene.
Geographical distribution; Theses species have widely distributed in the low to Middle Latitudes.
Order Foraminiferida (Eichwald, 1930)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Globigerinidae(Carpenter, Parker and Jones, 1862)
6- Parasubbotina varianata? (Olsson et al. )
Age is restricted from The Paleocene to lowermost Eocene
Stratigraphical Range Zone P1c to Zone E10.
Geographical distribution; Theses species have widely distributed in the high to low latitudes.
Order Foraminiferida (Eichwald, 1830)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Hedbergellidae (Loeblich and Tappan, 1961)
7- Turborotalia pomeroli [9]
Age Lower Eocene to Oligocene.
Stratigraphic Range Zone E10 to Oligocene.
Geographical distribution; Theses species have widely distributed in the Middle Latitudes.
Order Foraminiferida (Eichwald, 1830)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Hedbergellidae (Loeblich and Tappan, 1961)
8- Turborotalia frontosa [8]
Age Lower to Middle Eocene
Stratigraphical Range Zone E7 to Zone E11.
Geographical distribution; Theses species have world distributed.
Order Foraminiferida (Eichwald, 1930)
Super Family Globigerinaceae (Carpenter, Parker and Jones, 1862)
Family Globigerinidae(Carpenter, Parker and Jones, 1862)
9- Paragloborotalia (Turborotalia) griffinoides [6]
Age Eocene
Stratigraphical Range Zone E1 to Zone E16.
Geographical distribution; Theses species have widely distributed in the high latitudes.
B-Benthic Foraminifera
Class Foraminifera incertae sedis Order Lagenida
Superfamily Nodosarioidea Family Lagenidae
1- Lagena sp., (Walker and Boys, 1784) Age Eocene age
Geographical distribution; Theses species have world distributed.
Sub Order Rotaliina
Super Family Nummulitacea
Family Nummulitidae (de Blainville, 1827)
Subfamily Heterostegininae (Galloway, 1933)
2- Heterostegina sp., (Orbigny, 1826)
Age Eocene to Holocene
Geographical distribution; Theses species have world distributed.
Class Globothalamea
Order Rotaliida
Superfamily Planorbulinoidea
Family Cibicididae (Cushman, 1927)
Superfamily Planorbulinoidea Cushman, 1927
3-Cibicidoides micrus [10-29]
Age Early Eocene
Geographical distribution; Theses species have world distributed.
Class Globothalamea
Order Rotaliida
Family Alabaminidae Hofker, 1951
Genus Alabamina Toulmin, 1941
4- Al abamina sp.,
Age Eocene
Geographical distribution; Theses species have world distributed.
Class Globothalamea (Gumbel, 1870)
Order Rotaliida
Superfamily Nummulitoidea
Genus Discocyclina
5- Discocyclina (Discocyclina) marginata?, (Cushman, 1919)
Age Middle Eocene.
Geographical distribution; Theses species have distributed in middle latitude.
Conclusion
The availability of food and oxygen were the two main elements affecting the foraminiferal distribution. The number of species of (benthic, planktonic) foraminifera was directly impacted by the low oxygen levels caused by sediment intake at the time. It is revealed by the temporal decline in the proportion of infaunal benthic foraminifera that there was a brief period of reduced bottom-water oxygenation and nutrient availability (associated with the tectonic activities in the study area). The Paleocene-Eocene biostratigraphy of the area was examined in the Burton Guyot limestone column. The biostratigraphy of planktonic foraminifera in Burton Guyot limestone is complete, and this section’s age is based on overlapping ranges from the E10 (Luteian) to Middle Eocene. Nine planktonic foraminiferal biozones have been identified there.
References
- Expedition 330 Scientists (2012) Site U1376. In Koppers AAP, Yamazaki T, Geldmacher J and the Expedition 330 Scientists, Proe. IODP, 330: Tokyo (Integrated Ocean Drilling Program Management International, Inc.
- Gradstein FM, Ogg JG, Schmitz M, Ogg, G (2012) The Geologic Time Scale2012. Elsevier 2.
- Gradstein FM (2020) Chapter 3 – Evolution and Biostratigraphy, Eds: Felix M. Gradstein, James G. Ogg, Mark D. Schmitz, Gabi M. Ogg, Geologic Time Scale 2020. Elsevier Pg: 35-137.
- Bryan JR, Carter BD, Fluegeman RH, Krumm DK, Stemann TA (1997) The Salt Mountain Limestone of Alabama. Tulane Studies in Geology and Paleontology. 30: 1-60.
- Stainforth RM, Lamb JL, Luterbacher HP, Beard JH, Jeffords RM (1975) Cenozoic planktonic foraminiferal zonation and characteristics of index forms. Paleontological Contributions Article 62. P. 425 Smith WHF, Sandwell DT, 1997. Global sea floor topography from satellite altimetry and ship depth soundings. Science, 277: 1956-1962.
- Pearson PN, Olsson RK, Hemleben C, Huber BT, BerggrenWA (2006) Atlas of Eocene Planktonic Foraminifera. P. 1-513.
- Bolli HM (1957) Planktonic Foraminifera from the Eocene Navet and San Fernando formations of Trinidad. W.I. Bull. U.S. natl. Mus. 215: 155-172.
- Subbotina NN (1953) Iskopaemye foraminifery SSSR (Globigerinidy, Khantkenininidy i Globorotaliidy). Trudy Vsesoyznogo Nauchno-Issledovatel’skogo Geologo-razvedochnogo Instituta (VNIGRI) 76. P: 296.
- Toumarkine M, Bolli HM (1970) Evolution de Globorotalia cerroazulensis (Cole) dans l’Eocene moyen et superieur de Possagno (Italie). Revue de Micropaleontologie 13: 131-145.
- Bermudez PJ (1961) Contribucion al estudio de las Globigerinidea de la region Caribe-Antillana (Paleocene-Reciente). In: Boletino Geologia (Venezuela), Special Publicacion Vol. P. 22.
- Blow WH (1979) The Cainozoic Globigerinida. 3 Vols. P. 12.
- Jenkins DG (1971) New Zealand Cenozoic Planktonic Foraminifera. New Zealand Geological Survey Paleontological Bulletin 42.
- Kell JP, Watkins DK (2014) Data report: Calcareous nannofossil biostratigraphy of Expedition 330, Louisville Seamount Trail, Sites U1372-1377. In Koppers AAP, Yamazaki T, Geldmacher J and the Expedition 330 Scientists, Proe.IODP, 330: Tokyo (Integrated Ocean Drilling Program Management International, Inc.
- Khameiss B, Fluegeman R, Grigsby J, Malone S, Bernot R, et al. (2017) Biofacies Analysis of the Tertiary algal reef (Pinnacle Reef – Middle Eocene), IIA (Limestone Unit) At Burton Guyot (IODP Site U1376). 2017 -AIPG National Annual Conference. P. 23.
- Khameiss B, Fluegeman R (2017) Eocene Foraminifera from an Algal Reef Limestone at Burton Guyot (IODP site U1376), in the Southwest Pacific Geologic Problem Solving with Microfossils Conference. Microfossils IV. pg: 83-84.
- Khameiss B, Richard F (2017) Benthic Foraminifeal Paleoecology of Depositional Sequence TE 3.2 (Eocene; Bartonian) of the Eastern Gulf Coastal Plain. Geologic Problem Solving with Microfossils Conference. Microfossils IV.P. 67-68.
- Krasheninnikov VA, Basov IA (1983) Cenozoic planktonic foraminifers of the Falkland Plateau and Argentine Basin, Deep Sea Drilling Projejt Leg 71. In: Initial Reports of the Deep Sea Drilling Project Vol. 71 Eds: Ludwig WJ, Krasheninninov VA, P. 821-845.
- Lourens L, Sluijs A, Kroon D, Zchos Jc, Thomas E et al. (2005) Astronomical pacing of late Paleocene to early Eocene global warming events. Nature 435: 1083-1087.
- Pearson PN, Shackleton NJ, Hall MA (1993) The Stable Isotope Paleoecology of Middle Eocene Planktonic Foraminifera and Multi-species integrated Isotope Stratigraphy. Journal of Foraminiferal Research 23: 123-140.
- Poag CW, Commeau,JA (1995) Paleocene to middle Miocene planktic foraminifera of the southwestern Salisbury Embayment, Virginia and Maryland: biostratigraphy, allostratigraphy, and sequence stratigraphy. Journal of Foraminiferal Research 25: 134-155.
- Poore RZ, Bybell L (1988) Eocene to Miocene biostratigraphy of New Jersey Core ACGS # 4: Implications for regional stratigraphy. S. Geological Survey Bulletin 1829. P. 1-22.
- Poore RZ, Brabb EE (1977) Eocene and Oligocene planktonic foraminifera from the Upper Butano Sandstone and type San Lorenzo Formation, Santa Cruz Mountains, California. Journal of Foraminiferal Research 7: 249-272.
- Samanta BK (1970) Middle Eocene Planktonic Foraminifera from Lakhpat, Cutch, Western India. Micropaleontology 16.
- Spezzaferri S, Silva IS (1990) Oligocene planktonic foraminiferal biostratigraphy and paleoclimatic interpretation from Hole 538A, DSDP Leg 77, Gulf of Mexico. Paleogeography, Palaeoclimatology, Palaeoecology 83: 217-263.
- Snyder SW, Waters VJ (1985) Cenozoic planktonic foraminiferal biostratigraphy of the Goban Spur region. In: Deep Sea Drilling Project Vol. 80. P.23.
- Toumarkine M (1975) Middle and Late Eocene planktonic foraminifera from the northwestern Pacific, Leg 32 of the Deep-Sea Drilling Project. In: Initial Reports of the Deep Sea Drilling Project Vol. 32. eds: Larson RL, Moberly R pg: 735-751.
- Toumarkine M (1978) Planktonic Foraminiferal Biostratigraphy of the Paleogene of Sites 360 to 364 and the Neogene of Sites 362A, 363, and 364 Leg 40. In: Initial Results of the Deep-Sea Drilling Project Vol. 40.
- Toumarkine M, Luterbacher HP (1985) Paleocene and Eocene Planktonic Foraminifera. In: Plankton Stratigraphy. Pg: 87-154.
- Van Eijden AJM, Smit J (1991) Eastern Indian Ocean Cretaceous and Paleogene quantitative biostratigraphy. In: Proceedings of the Ocean Drilling Program, Scientific Results Vol. 121 Eds: Weissel, J. Peirce, J. Taylor, E. Alt, J. Pg: 77-123.