DOI: 10.31038/MIP.2024511
Abstract
Communities of enteropathogenic microorganisms adhere as biofilms to both natural and artificial surfaces encountered by eggs and chickens during production, constituting a major source of food cross-contamination. Given the rising bacterial resistance to chemical sanitary agents and antibiotics, there is a need to explore alternative approaches, particularly using natural products, to control the proliferation of these microorganisms along the surfaces of the poultry production chain. This study investigates and compares the bactericidal and antibiofilm properties of acetic, citric, and lactic acids against Salmonella enterica serovar Typhimurium and Escherichia coli O157 cells. Biofilms were allowed to develop on eggshells, stainless steel, and polystyrene surfaces at temperatures of 22°C and 37°C, and subsequently exposed to the acids for durations of 2 and 24 hours. The three organic acids exhibited varying degrees of reduction in planktonic, swarmer, and biofilm cells. Notably, acetic acid consistently produced the most promising outcomes, resulting in a reduction between 3 and 6.6 Log10 in the quantities of young and mature biofilm cells adhered to eggshells or stainless steel. Additionally, a decrease of 1 and 2.5 optical density units was observed in biofilms formed on the polystyrene surface. Overall, these findings suggest that acetic acid can effectively act as an anti-biofilm agent, disrupting both newly formed and matured biofilms formed under conditions encountered along the production chain of eggs and broilers.
Keywords
Food-contamination, Bactericidal, Organic acids, Enteropathogenic bacteria, Poultry production
Introduction
Foodborne pathogens, such as Salmonella enterica Serovar Typhimurium (S. Typhimurium) and E. coli O157, linked to poultry production and the food industry, are major concerns in global gastroenteritis outbreaks affecting humans. According to the USA Centers for Disease Control and Prevention, these pathogens contribute to 76 million infections, 325.000 hospitalizations, and 5.000 deaths annually in the USA alone [1]. In Colombia, a South American country, the Colombian National Institute of Health-Sivigila reported a total of 9.781 cases of foodborne illnesses involving 679 outbreaks in 2017 [2]. Despite the inherent protective physical and chemical barriers in eggs, research reveals that S. Typhimurium, E. coli O157, and other enteropathogenic bacteria can contaminate and infect them. Eggs typically become contaminated through three general routes: before oviposition, when the reproductive organs suffer an infection, and secondly, by encountering feces or contaminated surfaces [3,4]. Accumulating evidence illustrates that S. Typhimurium and E. coli O157, through the formation of biofilms, not only colonize eggs but also surfaces throughout the production chain (Figure. 1). This contamination of surfaces may result in the transmission of these pathogens, posing significant risks to public health [4-6].
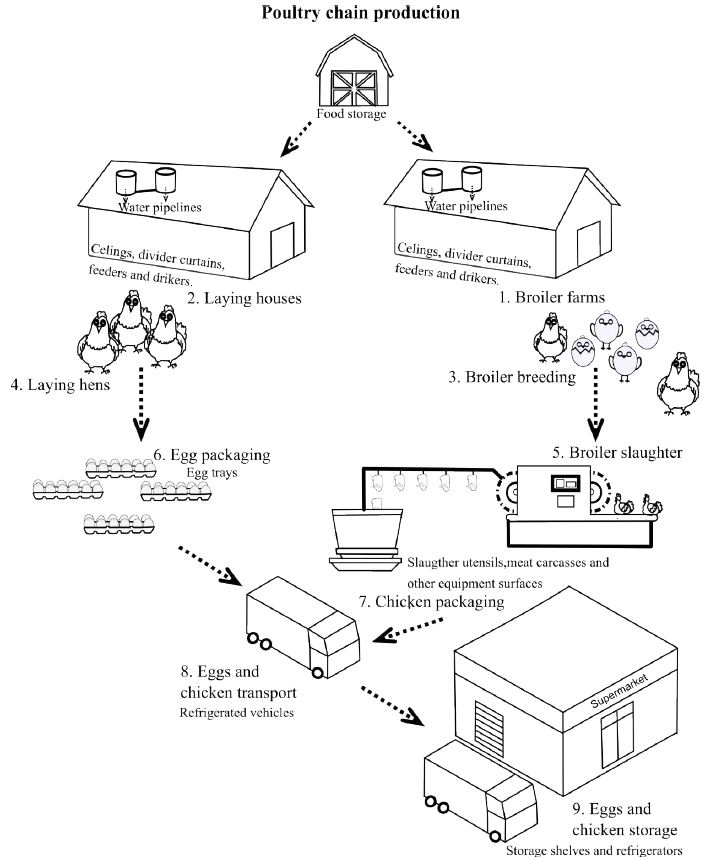
Figure 1: Areas and surfaces of the Colombian poultry production chain at risk for contamination by biofilms formed by enteropathogenic bacteria. Numbers highlight the different steps at which eggs and chickens can be contaminated by enteropathogenic bacteria. Italic letters indicate the places or utensils that may be made of stainless steel or polystyrene from which cross-contamination of eggs and chickens can occur.
Several studies have demonstrated how Salmonella and E. coli strains that are common causes of human gastroenteritis presented firm attachment of their bacterial cells to the eggshell surface and several types of foods and plants of production surfaces, facilitating the formation of biofilms [7-9]. The formation of a biofilm comprises several distinct steps. First, the initial reversible adsorption of cells onto the surface. Second, production of surface polysaccharides or capsular material occurs followed by the formation of an extracellular polymeric matrix. At this stage, biofilm cells form a strong attachment to the surface. In the following steps, the biofilm architecture is developed and matured. The process ends with the liberation of single motile cells that disperse into the environment and initiate again the process [10]. Biofilm formation is known to be influenced by several environmental cues, such as as availability and concentration of nutrients, and the physicochemical parameters of the of the surrounding environment, such as temperature and the material composition of the surface [11]. The surface type can influence microbial interactions among pathogens and promote co-biofilm formation, increases in individual pathogen biomass, and cell activity [12]. By nature, biofilm structure allows microbes to resist chemical or biological sanitizers, while bacterial cells are more vulnerable during the planktonic state and in a short contact time than when sequestered and protected in biofilms. Bacteria cells within biofilms are more resistant to environmental stresses, such as desiccation and UV light exposure, as well as to host-mediated responses, such as phagocytes [13]. Bacterial biofilms are more resistant to antimicrobial agents than are free-living cells, which makes it difficult to eradicate pathogens from surfaces commonly used in the poultry industry [5].
With the rise in the occurrence of foodborne outbreaks associated with poultry production, there is increasing interest in the use of novel biocide applications to prevent or reduce microbial contamination in food industries. The viability of microbes on food contact surfaces varies according to the biofilm state and formation ability, as well as the type of surface. Biofilm formation from the highest to lowest degree follows the order of eggshell > rubber > stainless steel > plastic [14,15]. As reported by Lee [15], rinsing surfaces with water, even extensively, appears to have limited effect on reducing S. Typhimurium biofilm viability. The regular application of cleaning and disinfecting procedures are common strategies employed to control pathogen establishment on industrial equipment [16]. Importantly, Chemical sanitizer efficacy can significantly depend on surface types, bacterial strains, and relative humidity [17]. Therefore, such procedures may not be fully effective in impeding or disrupting biofilms and can induce the formation and persistence of resistant phenotypes [18].
Novel alternatives, such as natural compounds extracted from bacterial cultures or aromatic plants, as well as organic acids, are currently under evaluation for their potential in eradicating biofilms. These compounds may exhibit high lethality against pathogens, efficiently penetrate the structure of a biofilm, and degrade easily in the environment [16]. Organic acids are generally recognized as safe (GRAS) by the USA Federal Drug Administration (FDA) and have been documented to possess antimicrobial activities against different pathogens [5]. In studies involving antibiotic-resistant bacteria, Clary [19] demonstrated how low concentrations (5%) of acetic acid rapidly killed (30 min) planktonic cells of Mycobacterium abscessus. On the other hand, Bradhan [20] demonstrated that lactic acid can decrease viable cell counts of planktonic as well as biofilm-forming cells of multiple carbapenem-hydrolyzing, multi-drug-resistant Klebsiella pneumoniae strains. Acetic acid demonstrated antimicrobial effectiveness on both smooth and rough cell morphotypes. Besides directly affecting bacterial cell viability, organic acids can also influence the electrochemical properties of the attachment surface, leading to an effective antimicrobial outcome [21].
An antimicrobial mechanism of organic acids, such as citric acid, acetic acid, and lactic acid, involves decreasing the environmental pH, creating unfavorable growth conditions for pathogenic bacteria [22]. Weak acids like acetic acid, when at a pH lower than their pKa and in their undissociated form, have shown the ability to reduce biofilm formation by permeating the biofilm structure and inner cell membrane. Kundukad [23] demonstrated that these weak acids, including acetic acid, could effectively eliminate bacteria without harming human cells if the pH remains close to their pKa. Organic acids in their undissociated form possess lipophilic properties, enabling them to diffuse across bacterial cell membranes, thereby disrupting cell function upon reaching the cell interior [5].
Research focusing on evaluating alternative treatments and methods to control S. Typhimurium and E. coli O157 biofilm formation on surfaces along the egg and other animal-derived food production chains is crucial to reduce cross-contamination. Accordingly, the present study aimed to assess the efficacy of organic acids in: 1) controlling biofilm formation by S. Typhimurium and E. coli O157 during the initial stages of development, and 2) disrupting mature biofilms. Eggshells, stainless steel, and polystyrene were utilized to simulate potential soiling surfaces encountered by eggs and broilers throughout the production chain. Two temperatures were assessed as key environmental variables: 22°C, representing the mean environmental temperature of the largest broiler-producing region in Colombia, and 37°C, simulating the optimal growth temperature of these pathogens. Additionally, to track the impact of exposure time and the potential development of resistance, the biofilms were subjected to organic acids for 2 and 24 hours.
Materials and Methods
Bacterial Strains and Growth Conditions
Bacterial strains used in this study were S. Typhimurium ATCC 14028 (American Type Culture Collection, Manassas, VA., USA) and E. coli O157 strain AGROSAVIA_CMSABV_Ec-col-B-001-2007 from the Animal Health collection of the AGROSAVIA Microbial Germplasm Bank (Mosquera, Cundinamarca, Colombia). The bacteria were grown on nutrient agar (Merck, Darmstadt, Germany) or Luria Bertani low salt agar (LBL) (peptone (ThermoFisher, Waltham, Massachusetts, USA) at 10 g.L-1, yeast extract (Merck) at 5 g.L-1, sodium chloride (Merck) at g.L-1, agar (Merck) at g.L-1. When required, LBL agar was acidified to pH 3 with 0.3% (v/v) acetic acid (Merck), 0.2% (v/v) citric acid (Merck), or 0.2% (v/v) lactic acid (Merck). For biofilm assays, LBL broth (LBL without agar) was used.
Growth Curves
S. Typhimurium and E. coli O157 were aerobically grown on LBL agar plates at 37°C for 24 h. The inocula were prepared by scraping the surface of the agar plates following the addition of 10 mL of LBL broth at pH 7 or acidified to pH 3 with acetic (0.3% v/v), citric (0.2% v/v) or lactic acid (0.2% v/v). These cell suspensions were adjusted to an OD at 600 nm of 0.1 (2.2 × 103 colony-forming units (cfu).mL-1) or 1.8 (2.8 × 109 103 cfu.mL-1). Three bacterial suspensions (n=3) per treatment and the control at an initial OD of 0.1 or 1.8 were incubated aerobically at 37°C for 48 h with constant shaking at 140 rpm. Every 2 h, 1-mL aliquots of the bacterial cultures were taken, and 10-fold serial dilutions and plating on LBL agar were made to determine Log10 cfu.mL-1 at each time.
Surface Spreading Assays
S. Typhimurium and E. coli O157 were grown aerobically in 5 mL of LBL broth at 37°C until reaching an optical density (OD) at 600 nm of 1 (16 h). Then, 1 mL of each culture was concentrated 10-fold by centrifugation at 4.400 × g for 5 min at room temperature. The pellets were suspended in 100 µL of LBL broth. The concentration of inocula for S. Typhimurium was 8.25 × 1010 and for E. coli O157 was 1.05 × 1011 cfu.mL-1. Semi-solid agar surface spreading plates were prepared as described by Amaya [24] with 20 mL of LBL broth containing 8% (w/v) of glucose and 0.6% (w/v) of agar and if required acidified with acid acetic (0.3% v/v), citric acid (0.2% v/v), or lactic acid (0.2% v/v). A 5-µL drop of the suspended bacteria was placed in the center of the plates (n=10) and allowed to air-dry for 10 min. The plates were inverted and incubated aerobically for 24 h at 37°C. The areas of the spreading colonies were measured with ImageJ software 1.52a (Wayne Rasband, National Institute of Health, Bethesda, MD, USA) by delimiting the coly area using the shaped and measured tools.
Disruption of Newly Formed Biofilms Developed on Eggshells and Stainless Steel
S. Typhimurium and E. coli O157 were grown aerobically on LBL agar plates at 37°C for 24 h. The inocula of the pathogens were prepared by scraping the cell mass grown from the surface of the plates and washing twice in 2 mL of LBL broth at pH 7 or LBL broth at pH 3, acidified with acid acetic (0.3% v/v), citric acid (0.2% v/v), or lactic acid (0.2% v/v), and centrifugation at 4.400 × g for 5 min at room temperature. Washed cells were suspended in 20 mL of the respective media. The OD of each suspension was adjusted to 1.8 at 600 nm (5 × 109 cfu.mL-1). Then, six 1-cm2 pieces of eggshell or stainless steel for each treatment, which were sterilized by autoclave at 15 lb of pressure and 121°C for 20 min, were weighted and covered with 5 mL of each bacterial suspension in 15-mL Falcon tubes. Negative controls contained each medium without inoculum. Following incubation for 2 or 24 h at 22°C or 37°C, eggshells and stainless-steel pieces were aseptically transferred with sterile forceps to 15-mL Falcon tubes. The eggshells and stainless-steel pieces were rinsed three times with 2 mL of sterile 0.85% NaCl solution to remove unbound cells. To detach the biofilm cells, the eggshells and stainless-steel pieces were sonicated twice in 2 mL of sterile 0.85% NaCl solution for 2 min with a pause of 2 min. Ten-fold serial dilutions were made in sterile 0.85% NaCl solution and plated using drop plate technique on nutrient agar. Plates were incubated aerobically at 37°C for 20 h and the numbers of colony-forming units were counted. The results were expressed as Log10 cfu.g-1 of eggshell or stainless steel.
Disruption of Mature Biofilms Formed on Eggshells and Stainless Steel
Pathogen biofilms were allowed to develop on the surface materials (n=9) for 2 or 24 h at 22 or 37°C in LBL broth (pH 7), following the procedures described above. Once the biofilms were formed, eggshells and stainless-steel pieces were aseptically transferred to LBL broth at pH 7 or acidified with acetic acid (0.3% v/v) to pH 3. The 2-h-old biofilms were incubated aerobically for 2 h and the 24-h-old biofilms were incubated for 24 h, at 22°C or 37°C. After rinsing three times with 2 mL of sterile 0.85% NaCl solution and sonication in 2 mL of sterile 0.85% NaCl solution, 10-fold serial dilutions were made and plated using drop plate technique on nutrient agar. Results were expressed as Log10 cfu.g-1 of eggshell or stainless steel.
Disruption of Biofilms Formed on Polystyrene
S. Typhimurium and E. coli O157 inocula were prepared as described above for the evaluation of biofilm formation on eggshells and stainless steel. To evaluate the disruption of young biofilms by acetic acid, ninety-six-well polystyrene plates (Becton Dickinson, Franklin Lakes, NY, USA) were inoculated with 180 µL of S. Typhimurium or E. coli O157 inoculum adjusted to an OD of 1.8 (approximately 5.12 × 109 cfu.mL-1) in LBL broth pH 7 or broth acidified to a pH of 3 with acetic acid (0.3% v/v), n=24. The multi-well plates were incubated aerobically for 2 or 24 h at 22 or 37°C, without shaking and under humid conditions to prevent evaporation. To evaluate the disruption of matured biofilms, first, the biofilms were allowed to form aerobically in LBL broth at pH 7 for 2 or 24 h. Subsequently, the culture broth was removed and 150 µL of LBL broth at pH 7 or broth acidified to pH 3 with acetic acid (0.3% v/v) was added to the wells, the number of wells used per treatment was of 24. Then the plates were incubated aerobically once more for an additional 2 or 24 h at 22°C or 37°C. Controls consisted of uninoculated broths. At the end of the incubation times, the OD was read at 600 nm using a SunriseTM microtiter plate reader (Tecan Group Ltd, Männedorf, Switzerland). Subsequently, the liquid contents of each well were gently removed, and the biofilms were stained for 1 h with 180 µL of 0.01% (w/v) crystal violet (Sigma-Aldrich, St. Louis, MO., USA) 3. Excess dye was removed, and the wells were rinsed three times with sterile distilled water. The plates were allowed to air-dry at room temperature before adding 180 µL of ethanol: acetone (80: 20) to each well. Crystal violet-stained biofilms were measured at 600 nm using a SunriseTM microtiter plate reader.
Statistical Analysis
At least three biological replicates of each experiment were carried out to ensure the reproducibility of results. Data of surface spread colony areas, cfu.g-1 of eggshell or stainless steel and crystal-violet stained biofilms were Log10 (x + 1) transformed to homogenize variances between treatments. Linear models (LM) were employed for statistical analyses using R v. 3.6.0 (http://www.R-project.org/) with packages lme4, car, and emmeans. Surface spreading data were analyzed using LM and pairwise comparisons were performed for the interaction between all factors. The cfu.g-1 of eggshell or stainless steel and OD data for multi-well plate assays were analyzed with a negative binomial distribution. The negative binomial theta parameter was established with an alternating iteration procedure using the glm.nb function. Pairwise multiple comparisons were carried out using the false discovery rate (FDR) for P-value corrections.
Results
Impact of Acetic Acid on S. Typhimurium and E. coli O157 Planktonic Cells
Growth curves were conducted to monitor the antimicrobial activity of the three organic acids on planktonic cells. Initial low (0.1 OD) and high (1.8 OD) concentrations of cells were employed to simulate the numbers used in young and mature biofilm inoculants, respectively. The results indicated that irrespective of the initial concentration (Figure 2C and 2D), all three organic acids exhibited bactericidal activity against S. Typhimurium and E. coli O157 planktonic cells. In both scenarios, a progressive decrease in colony-forming unit (cfu) numbers was observed over time. Compared to cultures at pH 7 with an initial OD of 0.1, cultures in LBL broth acidified with acetic, citric, and lactic acids exhibited reductions of 7.86 Log10 cfu.mL-1 for S. Typhimurium (Figure 2A) and 8.17 Log10 cfu.mL-1 for E. coli O157 (Figure 3B). When initial cell concentrations were high, cfu.mL-1 numbers also decreased in cultures acidified with the three organic acids. After 48 hours of incubation, viable Log cfu.mL-1 counts of S. Typhimurium and E. coli O157 in acidified cultures revealed reductions of 8.36 and 8.10, respectively.
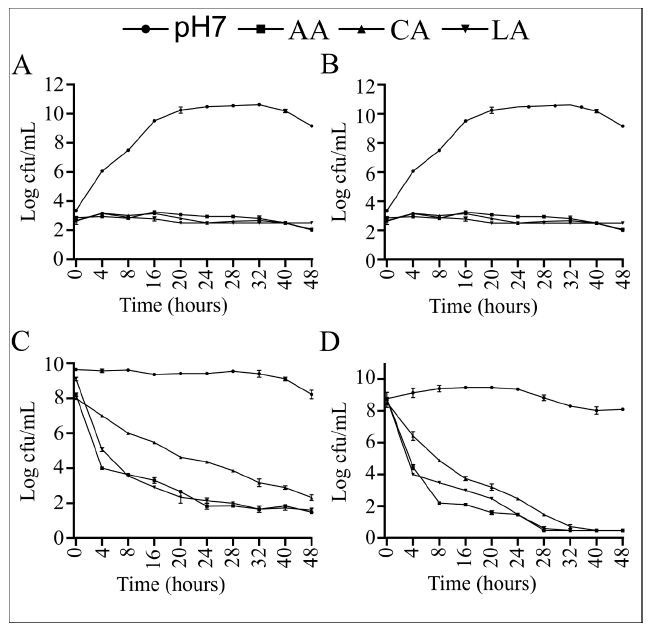
Figure 2: S. Typhimurium and E. coli O157 growth curves for control (pH 7) and acid (pH 3) broth cultures with an initial optical density of 0.1 (A and B, respectively) and 1.8 (C and D, respectively). Error bars indicate standard error of the mean (n=9).
Interference of Organic Acids with Surface Spreading
Bacterial surface motility is known to be involved at different stages of biofilm formation, especially at its initial stages. We evaluated the impact of acetic, citric, and lactic acids on this phenotype. Compared to control conditions, a significant (P < 0.05) decrease in the surface spreading abilities of S. Typhimurium and E. coli O157, ranging between 97 to 98%, was observed on the semi-solid agar plates containing any of the three organic acids (Figure 3).
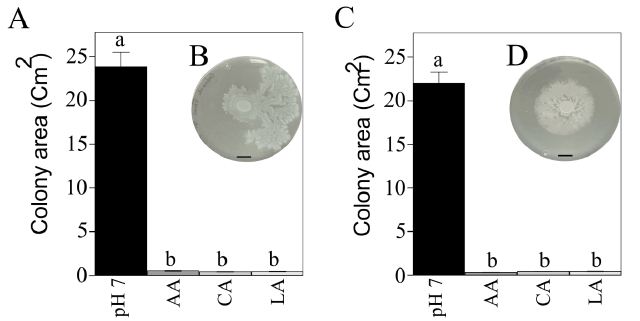
Figure 3: Effect of organic acids on S. Typhimurium (A) and E. coli O157 (C) surface spreading. Error bars indicate the standard error of the means (n=10). Bars with the same letter do not differ significantly (P > 0.05). B and D demonstrate the observed surface spreading patterns of S. Typhimurium and E. coli O157 at 24 h post-inoculation, respectively (bar=1 cm).
Disruption of Newly Formed Biofilms
First, the capacity of acetic, citric, and lactic acid to disrupt biofilms formed at 2 and 24 h post-inoculation (hpi) on eggshells was evaluated. Under the control treatment conditions, the numbers of S. Typhimurium and E. coli O157 attached cells were similar in most comparisons at 2 and 24 hpi (Table 1); although at 24 hpi at 37°C, fewer (P < 0.05) E. coli O157 than S. Typhimurium cells were found to be attached. Of the three acids, acetic acid generated (P < 0.05) higher reductions on newly formed biofilms developed by both pathogens, with an overall 3 Log10 cfu.g-1 of eggshells decrease at both times and temperatures compared to the controls. An exception was at 2 hpi and 37°C with biofilm formation by S. Typhimurium being controlled to a greater extent by lactic acid rather than by acetic and citric acids. Compared to the effect achieved by the other two organic acids, at 2 and 24 hpi, acetic acid also yielded the highest (P < 0.05) reduction of E. coli O157 biofilm formation at both temperatures.
Table 1: Organic acids inhibition of young Salmonella Typhimurium and Escherichia coli O157 biofilms developed on eggshells and stainless steel surfaces.
|
Bacteria (Incubation Temperature) |
||||||
|
Surface1 |
Treatment | Time (h) | ST (22°C) | ST (37°C) | EC (22°C) | EC (37°C) |
| ES | Con | 2 | 8.58 ± 0.05aA | 8.23 ± 0.12aA | 8.31 ± 0.11aA |
8.24 ± 0.13aA |
| 24 | 8.48 ± 0.06aA | 8.40 ± 0.18aA | 8.33 ± 0.09abA | 8.05 ± 0.10bA | ||
| AA | 2 | 5.12 ± 0.07bC | 7.36 ± 0.08aB | 5.03 ± 0.03aC |
5.12 ± 0.06bC |
|
| 24 | 5.13 ± 0.06aC | 5.07 ± 0.04aC | 5.03 ± 0.03aC | 5.06 ± 0.04aD | ||
| LA | 2 | 5.17 ± 0.08cB | 5.08 ± 0.05cC | 7.33 ± 0.29bB |
8.36 ± 0.06aA |
|
| 24 | 7.56 ± 0.21aB | 7.69 ± 0.07aB | 7.65 ± 0.06aB | 6.59 ± 0.07bC | ||
| CA | 2 | 6.33 ± 0.17bB | 8.13 ± 0.13aA | 7.52 ± 0.30aB |
6.23 ± 0.34bB |
|
| 24 | 7.59 ± 0.08cB | 8.14 ± 0.10Ba | 7.79 ± 0.05cB | 8.40 ± 0.11aB | ||
| SS | Con | 2 | 10.48 ± 0.01bA | 10.72 ± 0.01aA | 10.31 ± 0.01cA |
10.50 ± 0.01bA |
| 24 | 10.63 ± 0.01bA | 10.96 ± 0.00aA | 10.33 ± 0.01dA | 10.50 ± 0.01cA | ||
| AA | 2 | 4.04 ± 0.21aB | 4.23 ± 0.01aB | 4.20 ± 0.27aB |
4.23 ± 0.03aB |
|
| 24 | 4.41 ± 0.05bB | 4.45 ± 0.03bB | 4.61 ± 0.01aB |
4.59 ± 0.02aB |
||
1ES: Egg Shell, SS: Stainless Steel, Con: Control medium at pH 7, AC: Acetic acid medium at pH 3, LA: Lactic acid medium at pH 3, CA: Citric acid medium at pH 3, ST: Salmonella Typhimurium, EC: Escherichia coli O157.
abcdMeans (Log10 cfu/g) ± SE (n=9) in rows and with different letters are significantly different (P < 0.05).
ABCDMeans (Log10 cfu/g) ± SE (n=9) in columns, with the same surface material, and the same time, and with different letters are significantly different (P < 0.05).
The disruptive activity that citric and lactic acid caused on newly formed biofilms was found to depend on the time of exposure and the incubation temperature. Citric acid was found to be more effective in disrupting the 2-h-old biofilms formed by S. Typhimurium at 22°C, and by E. coli O157 at 37°C, causing a reduction in the number of cfu.g-1 of eggshell of 2.25 and 2.01 Log10, respectively. On the other hand, lactic acid exerted the higher antibiofilm activity against S. Typhimurium biofilms generating a decrease in the number of cfu attached per gram of eggshell at 22°C of 3.41 Log10 and at 37°C of 3.15 Log10. At the same time and temperatures, E. coli O157 biofilms saw a decrease of 1 and 0 Log10. Biofilms formed during 24 h, treated with this organic acid showed an overall decrease of less than 1 Log10 cfu.cm2-1 of eggshell, in both pathogens. Because acetic acid was observed to be the most effective organic acid in controlling S. Typhimurium and E. coli O157 biofilm formation on eggshells, this organic acid was selected for further studies.
As seen with eggshells, the number of cfu.g-1 of stainless steel attached at 2 and 24 h showed similar numbers by S. Typhimurium and E. coli O157 within control and acetic acid treatments (Table 1). The Attached S. Typhimurium cells to this surface were higher in the control treatment at 2 and 24 hpi (P < 0.05) at 37°C and lower (P < 0.05) for E. coli O157 at 22°C, although the differences were small. At all times and temperatures, acetic acid caused a reduction (P < 0.05) by nearly 6 Log10 of S. Typhimurium and E. coli O157 cfu.g-1 of stainless steel. All counts for acetic acid-treated biofilms were similar (P > 0.05) at 2 hpi; however, at 24 hpi, E. coli O157 counts at both temperatures were slightly higher (P < 0.05) than those for S. Typhimurium.
The formation of biofilms by both pathogens on multi-well polystyrene plates was also found to be influenced by temperature and incubation temperature (P < 0.05, Table 2). Lower (P < 0.05) biofilm OD values were found for S. Typhimurium and E. coli O157 at 22°C than at 37°C at 2 hpi and 24 hpi for the control and acetic acid treatments. Treatment with acetic acid resulted in both pathogens producing less (P < 0.05) biofilm at both temperatures when compared to control OD values at 2 and 24 hpi. However, there were higher decreases in OD values for both acetic acid-treated pathogens at both temperatures at 24 hpi as compared to 2 hpi. While an overall reduction of nearly 1 OD unit was obtained at 2 hpi for both pathogens, a decrease at 24 hpi of 2 OD units and 1.7 OD units was found for S. Typhimurium and E. coli O157, respectively.
Table 2: Acetic acid inhibition of young Salmonella Typhimurium and Escherichia coli O157 biofilms developed on polystyrene surfaces.
|
Bacteria (Incubation Temperature) |
||||||
|
Surface1 |
Treatment | Time (h) | ST (22°C) | ST (37°C) | EC (22°C) | EC (37°C) |
| PS | Con | 2 | 2.76 ± 0.04bA | 3.14± 0.03aA | 2.43 ± 0.07cA |
2.98 ± 0.05aA |
| 24 | 3.04 ± 0.02bA | 3.62 ± 0.09aA | 3.03 ± 0.03bA | 3.12 ± 0.04bA | ||
| AA | 2 | 1.68 ± 0.06bB | 1.84 ± 0.08abB | 1.68 ± 0.05bB |
1.91 ± 0.05aB |
|
|
24 |
0.64 ± 0.05cB | 1.59 ± 0.10aB | 1.22 ± 0.04bB |
1.53 ± 0.09aB |
||
1PS: Polystyrene, Con: Control medium at pH 7, AC: Acetic acid medium at pH 3, ST: Salmonella Typhimurium, EC: Escherichia coli O157
abcdMeans (OD) ± SE (n=24) in rows and with different letters are significantly different (P < 0.05).
ABMeans (OD) ± SE (n=24) in columns and with different letters are significantly different (P < 0.05).
Acetic Acid Disruption of Mature Biofilms
Control treatments showed that the number of cfu of S. Typhimurium and E. coli O157 attached per g of eggshell did not significantly increase from 2 to 24 h (P < 0.05) at any of the evaluated temperatures. On the other hand, 2 more hours of incubation were enough to allow higher (P < 0.05) numbers of S. Typhimurium and E. coli O157 cells to be attached to stainless steel than to eggshells for control and acetic acid-treated cultures at both temperatures. As observed in the assays of young biofilms, treatment with acetic acid for 2 and 24 h also generated significant (P < 0.05) reduction on the already formed and mature S. Typhimurium and E. coli O157 biofilms, regardless of the evaluated surface (Tables 3). Compared to control treatments, there was an overall 6.6 Log10 reduction in the number of cfu attached to eggshells and stainless-steel surfaces. Exposure to acetic acid for 2 h was enough to disrupt the already formed biofilms. Interestingly, prolonged exposure to acetic acid for 24 h did not incrementally affect these mature biofilms (Table 3). Furthermore, as observed when evaluating the disruption of young biofilms, the antibiofilm activity of acetic was higher on the biofilms formed on stainless steel than on eggshells.
Table 3: Acetic acid disruption of matured Salmonella Typhimurium and Escherichia coli O157 biofilms developed on eggshells and stainless surfaces.
|
Bacteria (Incubation Temperature) |
||||||
|
Surface1 |
Treatment | Time (h) | ST (22°C) | ST (37°C) | EC (22°C) | EC (37°C) |
| ES | Con | 2 | 8.67 ± 0.03aA | 8.77 ± 0.03aA | 8.31 ± 0.03bA |
8.32 ± 0.05bA |
| 24 | 8.78 ± 0.08bA | 8.94 ± 0.01aA | 8.39 ± 0.03cA | 8.33 ± 0.03cA | ||
| AA | 2 | 2.24 ± 0.01aB | 2.23 ± 0.01aB | 2.21 ± 0.02aB |
2.22 ± 0.03aB |
|
| 24 | 2.41 ± 0.05bB | 2.45 ± 0.03bB | 2.59 ± 0.01aB | 2.59 ± 0.02aB | ||
| SS | Con | 2 | 10.54 ± 0.02bA | 10.67 ± 0.02aA | 10.11 ± 0.01cA |
10.68 ± 0.02aA |
| 24 | 10.76 ± 0.04bA | 10.92 ± 0.01aA | 10.85 ± 0.01aA | 10.92 ± 0.00aA | ||
| AA | 2 | 3.46 ± 0.03bB | 3.65 ± 0.01aB | 3.57 ± 0.03aB |
3.62 ± 0.04aB |
|
|
24 |
3.56 ± 0.04cB | 3.88 ± 0.02aB | 3.55 ± 0.07cB |
3.72 ± 0.04bB |
||
1ES: Egg Shell, SS: Stainless Steel, PS: Polystyrene, Con: Control medium at pH 7, AC: Acetic acid medium at pH 3, LA: Lactic acid medium at pH 3, CA: Citric acid medium at pH 3, ST: Salmonella Typhimurium, EC: Escherichia coli O157
abcdMeans (Log10 cfu/g) ± SE (n=9) in rows and with different letters are significantly different (P < 0.05).
ABCDMeans (Log10 cfu/g) ± SE (n=9) in columns, with the same surface material, and with different letters are significantly different (P < 0.05).
Incubation of the mature biofilms formed on the polystyrene surface for an additional 2 and 24 h generated significant differences (P < 0.05) in the OD values for S. Typhimurium and E. coli O157 biofilms. Regardless of the time and temperature of incubation, the OD values of E. coli O157 biofilms were higher than those of S. Typhimurium. Additionally, while S. Typhimurium showed higher OD values at 24 hpi than at 2 hpi, E. coli O157 OD values were reduced over time. An overall reduction on the OD values caused by acetic acid was observed in the matured biofilms formed by the two pathogens, however the antibiofilm activity of the acid varied depending on the time and temperature (P < 0.05). The higher antibiofilm activity exerted by acetic acid on S. Typhimurium matured biofilms was observed at 24 hpi and 22°C (1.08). Similarly, the higher reduction in the OD values in E. coli O157 was found at 22°C; although it was observed at 2 (2.49) and 24 hpi (2.33). Extending the exposure to acetic acid of S. Typhimurium matured biofilms formed at 22°C led to a higher reduction on the OD values at 24 hpi than at 2 hpi. However, this decrease caused by a longer exposure to acetic acid was not observed for the matured biofilms formed at 37°C by S. Typhimurium or by E. coli O157 at any of the evaluated temperatures (Table 4).
Table 4: Acetic acid disruption of matured Salmonella Typhimurium and Escherichia coli O157 biofilms developed polystyrene surfaces.
|
Bacteria (Incubation Temperature) |
||||||
|
Surface1 |
Treatment | Time (h) | ST (22°C) | ST (37°C) | EC (22°C) | EC (37°C) |
| PS | Con | 2 | 2.60 ± 0.05bA | 1.19 ± 0.08cA | 4.40 ± 0.06aA |
2.77 ± 0.07bA |
|
24 |
3.57 ± 0.06bA |
2.09 ± 0.06dA | 4.20 ± 0.08aA |
2.35 ± 0.08cA |
||
|
AA |
2 |
2.20 ± 0.15aB | 0.46 ± 0.07bB | 1.91 ± 0.1aB |
1.75 ± 0.09aB |
|
|
24 |
2.49 ± 0.11aB | 1.32 ± 0.14cB | 1.87 ± 0.08bB |
1.53 ± 0.11abB |
||
1PS: Polystyrene, Con: Control medium at pH 7, AC: Acetic acid medium at pH 3, ST: Salmonella Typhimurium, EC: Escherichia coli O157
abcdMeans (OD) ± SE (n=24) in rows for each surface and with different letters are significantly different (P < 0.05).
ABMeans (OD) ± SE (n=9) in columns, with the same time, and with different letters are significantly different (P < 0.05).
Discussion
Complete removal of enteropathogenic bacteria from the poultry production chain environment is essential to ensure overall food safety. Pathogens like S. Typhymurium and E. coli O157 possess the capability to form biofilms, enabling their survival under unfavorable conditions by adhering to abiotic surfaces such as metals, plastic, or glass while creating a protective barrier [25,26]. Despite the implementation of numerous hygienic measures, concerns persist regarding the efficacy of disinfectants due to the emergence of bacterial resistance [27]. Moreover, several chemical sanitizers previously used for human health purposes are now prohibited, leading to a renewed interest in substituting chemical industrial sanitizers with natural antimicrobial agents. Organic acids, considered safe for food animal and human health, stand out as exceptional alternatives in this regard [28]. They are affordable and known to be safe compounds.
The results from the current study demonstrate the efficacy of acetic acid as an antibiofilm agent against S. Typhimurium and E. coli O157 biofilms. Halstead [29] similarly revealed the bactericidal actions of this organic acid against pathogens such as E. coli, Staphylococcus aureus, and Acinetobacter baumannii. However, in contrast to these findings, other studies have suggested that acetic acid might not be the most efficient biofilm disruptor when compared to other organic acids. For instance, Ban [30] evaluated the antibiofilm activities of propionic acid, acetic acid, lactic acid, malic acid, and citric acid, and found lactic acid to be the most effective in disrupting 6-day-old S. Typhimurium, E. coli O157: H7, and Listeria monocytogenes biofilms. Moreover, Amrutha [5] reported that, when comparing the activity of acetic, lactic, and citric acids at a 2% concentration, lactic acid achieved maximum inhibition of Salmonella sp. and E. coli biofilms formed on cucumber. The degree of antimicrobial effect might be influenced by the concentration of organic acid and the exposure time [28]. According to Beier [31], acetic, butyric, and propionic acids required lower molar amounts than citric, formic, and lactic acids to significantly inhibit enteropathogens. Furthermore, Bardhan [20] indicated that lactic acid was an effective antimicrobial against clinical carbapenem-hydrolyzing, multi-drug-resistant Klebsiella pneumoniae planktonic and biofilm-forming cells. The authors observed cell membrane damage and high rates of bacteriolysis after treatment with lactic acid at concentrations of 0.15% and 0.225%.
The antibacterial activity of organic acids has been associated with their pKa and the optimal pH for dissociation [28]. The pKa values for acetic, citric, and lactic acid are 4.476, 3.86, and 3.13, respectively. Kundukad [23] demonstrated that maintaining their pH close to their pKa enables weak acids like acetic and citric acid to eliminate persistent cells within biofilms of antibiotic-resistant bacteria such as Klebsiella pneumoniae KP1, Pseudomonas putida OUS82, Staphylococcus aureus 15981, Pseudomonas aeruginosa DK1-NH57388A, and P. aeruginosa PA_D25. When provided at a pH lower than their pKa, these compounds can penetrate the biofilm matrix and bacterial cell membranes. While lactic acid is considered a stronger acid than acetic acid based on their pKa values, the efficacy of organic acids also relies on pH levels. The proximity between the pKa value of lactic acid and the pH of 3 used in this study might explain why acetic acid exhibited better performance against biofilm formation and disruption than lactic acid. Further studies comparing the effectiveness of these organic acids at various pH values are necessary to confirm these observations.
In general, it has been suggested that increasing the contact time with disinfectants enhances their antibiofilm activities on various material surfaces [15]. In the current study, it was observed that prolonged exposure of S. Typhimurium and E. coli O157 planktonic cells to the tested organic acids resulted in lower OD values, as depicted by the presented growth curves. However, when mature biofilms of these microbes were exposed to acetic acid on polystyrene surfaces, this time-related effect was not observed. The OD values for mature biofilms did not decrease after exposure to acetic acid for 2 or 24 hours. Similar resistance over time was noted for biofilm cells attached to eggshells and stainless steel when the biofilm formation and contact time with organic acids extended from 2 hours to 24 hours. Amrutha [5] reported that exposure to acetic, citric, and lactic acids did not significantly reduce the production of exopolysaccharides in Salmonella sp. biofilms and resulted in reductions of 10.89%, 6.25%, and 13.42% in E. coli O157: H7 biofilms, respectively. The extracellular matrix developed by biofilm cells acts as a barrier, impeding the penetration or inactivation of antimicrobial compounds [31,32]. Therefore, the limited reduction in S. Typhimurium and E. coli O157 biofilms formed on eggshells, stainless steel, and polystyrene with increased exposure time to acetic acid is likely due to the obstruction presented by the biofilm matrix against the passage of organic acids. Research focusing on disrupting the biofilm matrix using alternative methods before exposure to organic acids could lead to the development of complementary approaches to enhance the antimicrobial activity of organic acids.
In addition to the biofilm matrix defensive shield, it is conceivable that the remaining cells inside the S. Typhimurium and E. coli O157 biofilms would respond to the effects of acetic acid by triggering other protection strategies. Changes in membrane lipids have been described as one of these defensive mechanisms [33]. Additional cell protective strategies would include the release of ammonia [34], the pumping out of protons, and the proton-consuming decarboxylation processes. More recently, Clary [19] demonstrated how the bacterial colony diversification (morphotype) would define the outcome of tolerance to a particular stressor during the process of biofilm formation and its persistence against environmental assaults. Amrutha [5] demonstrated that a reduction of exopolysaccharide (EPS) synthesis, EPS composition and organization, swimming and swarming cell patterns, and a negative impact on quorum sensing play crucial roles in microbial community architecture as well as resistance to toxic substances. Further research is required to identify which of these mechanisms are used by the remaining S. Typhimurium and E. coli O157 biofilm cells attached to eggshells and the industrial surfaces evaluated in the current study.
The antibiofilm activity of organic acids, such as acetic acid, might encounter hindrances due to alterations in the biofilm structure caused by temperature shifts and variations in adhesion surface types. Generally, temperature and surface material have been reported to influence the attachment ability of enteropathogenic and other bacteria, consequently affecting the biofilm structure [35]. In the current study, we did not assess the impact of temperature on the biofilm structure on the tested surfaces. However, our findings indicate that at 22°C, acetic acid exhibited less control only over mature biofilms formed by S. Typhimurium on polystyrene, differing from the conditions at 37°C. Similar temperature-related alterations in biofilm capacity were observed by Andersen [36] when evaluating the biofilm-forming capacity of several E. coli K12 clinical isolates. They reported a higher number of attached cells at 30°C compared to 35°C, observing denser and more evenly distributed biofilms on silicone surfaces at the lower temperature. Andersen [36] suggested that the presence of curli fibers, which facilitate cell adhesion, might have influenced the type and creation of the biofilm structure, particularly at lower temperatures where these cell surface adhesins are produced. Furthermore, another study focused on E. coli O157: H7 biofilms formed at 4°C and 15°C on beef processing surfaces concluded that while a slight decrease in the number of attached cells was noted at 4°C, it did not hinder the overall increase in attached cell numbers over time [37].
Conclusion
The efficacy of compounds utilized for sanitation involves multifaceted events associated not only with the morphology and physiology of the target microbial cells but also with factors such as relative surface hydrophobicity, material surface roughness, and the impact of shear stress [38]. Organic acids can influence the internal chemical equilibrium of microbial cells, leading to alterations in cell membrane integrity or cellular activities, ultimately resulting in cell death. Consequently, organic acids represent an important option for sanitizing purposes and may potentially be combined or incorporated into innovative carrier matrices with other established antimicrobial molecules, such as essential oil components, thereby improving molecule stability and extending their biological activity [39]. The results obtained from this study offer new insights into the effectiveness of acetic acid as an antibiofilm agent, which can be utilized to control S. Typhimurium and E. coli O157 biofilms formed under conditions encountered along the poultry production chain. This newfound information may facilitate the integration of this natural compound into hygiene programs aimed at preventing cross-contamination of eggs, broilers, and broiler meat products.
Acknowledgments
This work supported by the United States Department of Agriculture under grant number 58-3091-7-028-F; and by the Colombian Ministry of Agriculture and Rural Development under grant numbers Tv18 and Tv19. We thank Yessica Muñoz and Xiomara Abella for technical assistance, and Corporación Colombiana de Investigación Agropecuaria – Agrosavia for supporting this research.
Contributions
AGC and CVAG were involved in the experimental and performed biofilm experiments. AGC, MEH, FRV and CVAG participated in data analysis and wrote the manuscript.
Ethics Approval
Not applicable.
Consent to Participate
All authors approved the manuscript.
Consent for Publication
The authors consented for the publication.
Statements and Declarations
Competing Interests
The authors declare no competing interests.
References
- Afzal A, Hussain A, Irfan M, and Malik KA (2015) Molecular diagnostics for foodborne pathogens (Salmonella spp.) from poultry. Life Sci 2: 91-97.
- Instituto Nacional de Salud de Colombia (2017) Investigación de brote enfermedades transmitidas por alimentos y vehiculizadas por agua, 59(2): 4-16.
- Gantois I, Ducatelle R, Pasmans F, Haesebrouck F, et al. (2004) Cross-sectional analysis of clinical and environmental isolates of Pseudomonas aeruginosa: biofilm formation, virulence, and genome diversity. Pharmacol 72: 133-144. [crossref]
- Pande VV, McWhorter AR, Chousalkar KK (2016) Salmonella enterica isolates from layer farm environments are able to form biofilm on eggshell surfaces. Biofouling 32: 699-710.
- Amrutha B, Sundar K, Halady Shetty PH (2017) Effect of organic acids on biofilm formation and quorum signaling of pathogens from fresh fruits and vegetables. Microb Pathog 111: 156-162. [crossref]
- Chowdhury MAH, Ashrafudoulla, Mevo SIU, Mizan MFR, et al. (2023) Current and future interventions for improving poultry health and poultry food safety and security: A comprehensive review. Compr Rev Food Sci Food Saf 22: 1555-1596. [crossref]
- Yang X, Tran F, Youssef MK, Gill CO (2015) Determination of sources of Escherichia coli on beef by multiple-locus variable-number tandem repeat analysis. J Food Prot 78: 1296-1302. [crossref]
- Silva PL, Goulart LR, Reis TF, Mendonça EP, Melo RT, et al. (2019) Biofilm formation in different Salmonella serotypes isolated from poultry. Curr Microbiol 76: 124-129. [crossref]
- Harrell JE, Hahn MM, D’Souza SJ, Vasicek EM, Sandala JL, et al. (2021) Salmonella biofilm formation, chronic infection, and immunity within the intestine and hepatobiliary tract. Front Cell Infect Microbiol 10: 624622. [crossref]
- Kim SH, Wei CI (2007) Biofilm formation by multidrug-resistant Salmonella enterica serotype Typhimurium phage type DT104 and other pathogens. J Food Prot 70: 22-29.
- Schonewille E, Nesse LL, Hauck R, Windhorst D, Hafez HM, Vestby LK (2012) Biofilm building capacity of Salmonella enterica strains from the poultry farm environment. FEMS Microbiol Immunol 65: 360-365. [crossref]
- Maggio F, Rossi C, Chaves-López C, Serio A, Valbonetti L, Pomilio F, et al. (2021) Interactions between Listeria monocytogenes and Psuedomonas fluorescens in dual-species biofilms under simulated dairy processing conditions. Foods 10: 176. [crossref]
- Fatemi P, Frank JF (1999) Inactivation of Listeria monocytogenes/Pseudomonas biofilms by peracid sanitizers. J Food Prot 62: 761-765.
- Hingston PA, Stea EC, Knøchel S, Hansen T (2013) Role of initial contamination levels, biofilm maturity and presence of salt and fat on desiccation survival of Listeria monocytogenes on stainless steel surfaces. Food Microbiol 36: 46-56. [crossref]
- Lee KH, Lee JY, Roy PK, Mizan MFR, Hossain MI, et al. (2020) Viability of Salmonella Typhimurium biofilms on major food-contact surfaces and eggshell treated during 35 days with and without water storage at room temperature. Poult Sci 99: 4558-4565. [crossref]
- Bridier A, Sanchez-Vizuete P, Guilbaud M, Piard JC, et al. (2015) Biofilm-associated persistence of food-borne pathogens. Food Microbiol 45: 167-178. [crossref]
- Joseph B, Otta SK, Karunasagar I, Karunasagar I (2001) Biofilm formation by Salmonella on food contact surfaces and their sensitivity to sanitizers. Int J Food Microbiol 64: 367-372. [crossref]
- Simoes M, Simoes LC, Vieira MJ (2010) A review of current and emergent biofilm control strategies. LWT-Food Sci Technol 43: 573-583.
- Clary G, Sasindran SJ, Nesbitt N, Mason L, Cole S, Azad A, McCoy K, Schlesinger LS, Hall-Stoodley L (2018) Mycobacterium abscessus smooth and rough morphotypes form antimicrobial-tolerant biofilm phenotypes but are killed by acetic acid. Antimicrob Agents Chemother 62: e01782-17. [crossref]
- Bardhan T, Chakraborty M, Bhattacharjee B (2019) Bactericidal activity of lactic acid against clinical, carbapenem-hydrolyzing, multi-drug-resistant Klebsiella pneumoniae planktonic and biofilm-forming cells. Antibiotics 8: 181. [crossref]
- Souza JG, Cordeiro JM, Lima CV, Barão VA (2019) Citric acid reduces oral biofilm and influences the electrochemical behavior of titanium: An in situ and in vitro study. J Periodontol 90(2): 149-158. [crossref]
- Canibe N, Steien SH, Øverland M, Jensen BB (2001) Effect of K-diformate in starter diets on acidity, microbiota, and the amount of organic acids in the digestive tract of pig. J Anim Sci 79: 2123-2133. [crossref]
- Kundukad B, Schussman M, Yang K, Seviour T, Yang L, et al. (2017) Mechanistic action of weak acid drugs on biofilms. Sci Rep 7: 1-12. [crossref]
- Amaya-Gómez CV, Porcel M, Mesa-Garriga L, Gómez-Álvarez MI (2020) A framework for the selection of plant growth-promoting rhizobacteria based on bacterial competence mechanisms. Appl Environ Microbiol 86: e00760-20. [crossref]
- Peng D (2016) Biofilm formation of Salmonella. Microbial Biofilms. Biofilms-Importance and Applications. IntechOpen, 231-242.
- Yang X, Wang H, Hrycauk S, Holman DB, Ells TC (2023) Microbial dynamics in mixed-culture biofilms of Salmonella Typhimurium and Escherichia coli O157: H7 and bacteria surviving sanitation of conveyor belts of meat processing plants. Microorganisms 11: 421. [crossref]
- Yuan L, Sadiq FA, Wang N, Yang Z, He G (2020) Recent advances in understanding the control of disinfectant-resistant biofilms by hurdle technology in the food industry. Crit Rev Food Sci Nutr 1-16. [crossref]
- Coban HB (2020) Organic acids as antimicrobial food agents: applications and microbial productions. Bioprocess Biosyst Eng 43: 569-591.
- Halstead FD, Rauf M, Moiemen NS, Bamford A, Wearn CM, et al. (2015) The antibacterial activity of acetic acid against biofilm-producing pathogens of relevance to burns patients. PLoS One 10: e0136190. [crossref]
- Ban GH, Park SH, Kim SO, Ryu S, Kang DH (2012) Synergistic effect of steam and lactic acid against Escherichia coli O157: H7, Salmonella Typhimurium, and Listeria monocytogenes biofilms on polyvinyl chloride and stainless steel. Int J Food Microbiol 157(2): 218-223. [crossref]
- Beier RC, Harvey RB, Hernandez CA, Hume ME, et al. (2018) Interactions of organic acids with Campylobacter coli from swine. PLoS One 13: e0202100.
- Kim SH, Wei CI (2007) Biofilm formation by multidrug-resistant Salmonella enterica serotype Typhimurium phage type DT104 and other pathogens. J Food Prot 70: 22-29.
- Pienaar JA, Singh A, Barnard TG (2020) Membrane modification as a survival mechanism through gastric fluid in non-acid adapted enteropathogenic Escherichia coli (EPEC). Microb Pathog 144: 104180. [crossref]
- Lu P, Ma D, Chen Y, Guo Y, et al. (2013) L-glutamine provides acid resistance for Escherichia coli through enzymatic release of ammonia. Cell Res 23: 635-644. [crossref]
- Lund P, Tramonti A, De Biase D (2014) Coping with low pH: Molecular strategies in neutralophilic bacteria. FEMS Microbiol Rev 38(6): 1091-1125.
- Andersen TE, Kingshott P, Palarasah Y, Benter M, et al. (2010) A flow chamber assay for quantitative evaluation of bacterial surface colonization used to investigate the influence of temperature and surface hydrophilicity on the biofilm-forming capacity of uropathogenic Escherichia coli. J Microbiol Methods 81: 135-140. [crossref]
- Dourou D, Beauchamp CS, Yoon Y, Geornaras I, Belk KE, et al. (2011) Attachment and biofilm formation by Escherichia coli O157: H7 at different temperatures, on various food-contact surfaces encountered in beef processing. Int J Food Microbiol 149: 262-268. [crossref]
- Cai S, Phinney DM, Heldman DR, Snyder AB (2020) All treatment parameters affect environmental surface sanitation efficacy, but their relative importance depends on the microbial target. Appl Environ Microbiol 87: e01748-20. [crossref]
- Scaffaro R, Lopresti F, Marino A, Nostro A (2018) Antimicrobial additives for poly (lactic acid) materials and their applications: current state and perspectives. Appl Microbiol Biotechnol [crossref]