Abstract
Most leaves in angiosperms have reticulate venation, but not all leaves with reticulate venation belong to angiosperms. Although angiosperms flowers have been reported in the Jurassic, leaves similar to angiosperms are lacking in the Jurassic. Here we report an unnamed fossil leaf from the Middle Jurassic Yan’an Formation of Ningxia, China. Although its partial preservation does not allow us to determine the affinity of the fossil, its occurrence underscores the probablity that future digging may uncover angiosperm leaves in the Jurassic.
Keywords
Fossil, Middle Jurassic, China, Angiosperms, Leaf
The origin and early evolution of angiosperms, which have more than 300,000 species and account for more than 90% species diversity of land plants, have been foci of botanical debates for long time. Some palaeobotanists thought that angiosperms did not occur on the Earth until the Cretaceous. But recent years witnessed increasing reports of angiosperms in the pre-Cretaceous age. Among the reports, Schmeissneria [1,2] and Nanjinganthus [3,4], both from the Early Jurassic, are based on tens even hundreds of specimens, making a strong case that angiosperms have long existed in the pre-Cretaceous. Theoretically, angiosperm leaves have more potential to be preserved as fossils. But the fact is that angiosperm leaves are almost completely lacking in the Jurassic. This situation makes any trace of angiosperm-like leaves in the Jurassic especially badly wanted. Here we report a partially preserved leaf from the Yan’an Formation (the Middle Jurassic) of Lingwu, Ningxia, China (37°43’N, 106°26’E, Figure 1). The character assemblage of reticulate venation and intramarginal vein of this leaf are only seen in angiosperms hitherto, making it unique in the fossil record. This discovery makes future discovery of angiosperm leaves in the Jurassic more likely.
Figure 1: Geographical information of the fossil locality in Lingwu, Ningxia, China (37˚43’N, 106˚26’E). a. Fossil locality (black Square) in northwestern China. b. Detailed position of fossil locality (black square) in Lingwu, Ningxia, China.
The specimen (No. SGY007-16, deposited in Ningxia Geological Museum) was preserved as a compression with some coaly residue, uncovered from the Yan’an Formation (the Middle Jurassic), which is widely distributed in Northeastern China and has yielded various fossil plants [5-8]. The specimen is a grey siltstone slab 36 mm x 26 mm. The details were imaged using a Nikon SMZ1500 stereomicroscope equipped with a Nikon DS-Fi1 digital camera. All figures are organized using a Photoshop 7.0.
The leaf is incomplete, at least 15 mm long, 4 mm wide (Figure 2a). The leaf is smooth-margined, with an intramarginal vein (Figure 2a, 2b and 2d). Lateral veins parallel each other, branching from the midvein at an angle between 40° and 50° (Figure 2a and 2b). Lateral veins and transverse veins in between form angular meshes, which are 0.44-1.22 mm long and 0.2-0.45 mm wide (Figure 2a-2c). Lateral and transverse veins form right angles or acute angles, about 88 μm wide, with no obvious differentiation between lateral and transverse veins (Figure 2a-2c). Rarely, there is a freely-ending veinlet in an areole (Figure 2c).
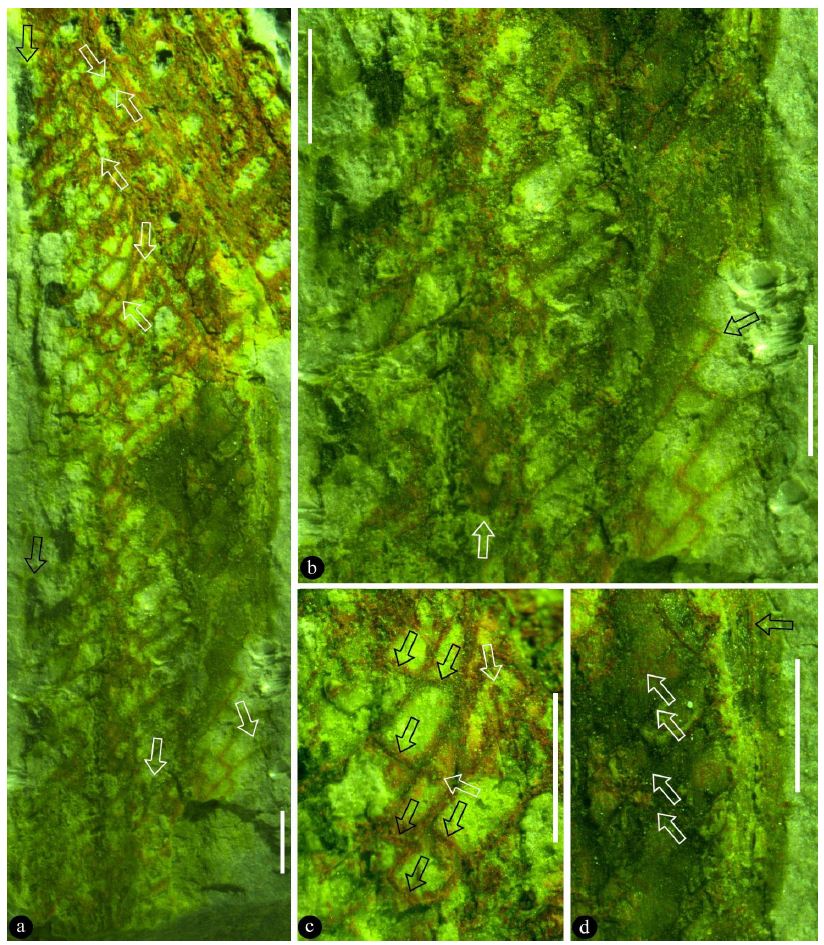
Figure 2: The partially preserved leaf and its details. All scale bar=1 mm. a. The general view of the specimen, showing the partial lamina and partially preserved midvein (black arrow) and the freely ending veinlets (white arrows). b. Detailed view of the basal portion of the leaf, showing a branch in the background (white arrow) and vein (black arrow) after skeletonization. c. Vein meshes after skeletonization, showing the transverse veins (black arrows) between lateral veins. Note the freely ending veinlet (white arrow). d. Detailed view showing smooth margin with intramarginal vein (black arrow) and parallel lateral veins (white arrows).
Other than in angiosperms, reticulate venation has been in several fossil taxa, including ferns (Clathropteridaceae, Dipteridaceae, Polypodiaceae), uncertain group (Gigantopteridales), seed plants (Glossopteridales, Caytoniales, Ginkgoales, Cycadales, Gnetales, angiosperms, uncertain groups) (Table 1). Therefore the occurrence of reticulate venation does not ensure that a taxon with reticulate venation is an angiosperms [9-12].
Table 1: Comparison of our leaf and previously reported taxa with reticulate venation. Note that intramarginal vein is restricted to angiosperms.
|
Affinity |
Margin |
Reticulate venation |
Intramarginal vein |
Vein order |
Midrib |
Secondary vein |
Freely ending veinlet |
Age |
Ref |
|
| Clathropteris | Clathropteridaceae | toothed | frequent | absent | 4+ | present | unbranched | absent | Mesozoic | |
| Hausmannia | Dipteridaceae | toothed | frequent | absent | 3 | absent | dichotomous | absent | Mesozoic | 9 |
| Polypodium | Polypodiaceae | toothed | frequent | absent | 3 | present | unbranched | absent | extant | 10 |
| Woodwardia | Polypodiaceae | smooth | frequent | absent | 3? | present | dichotomous | absent | extant | 10 |
| Onoclea | Polypodiaceae | smooth | frequent | absent | 2 | present | ? | absent | extant | 10 |
| Linopteris | Seed plants | smooth | frequent | absent | 1 | present? | dichotomous | absent | Palaeozoic | 10 |
| Reticulopteris | Seed plants | smooth | frequent | absent | 2 | present | dichotomous | absent | Palaeozoic | 10 |
| Lonchopteris | Seed plants | smooth | frequent | absent | 2 | present | dichotomous | absent | Palaeozoic | 10 |
| Ginkgo biloba | Ginkgoales | infrequent | 1 | absent | absent | extant | 10 | |||
| Stangeria | Cycadales | frequent | 2 | multi-strand | dichotomous | absent | extant | 10 | ||
| Ctenis | Cycadales | infrequent | 1 | absent | absent | Mesozic | 10 | |||
| Dictyozamites | Bennettitales | frequent | 1 | absent | absent | Mesozoic | 10 | |||
| Drewria | Gnetales | frequent | 2 | absent | unbranched | absent | Cretaceous | 10 | ||
| Welwitschia | Gnetales | frequent | 2 | absent | unbranched | absent | extant | 10 | ||
| Gnetum | Gnetales | frequent | 4 | multi-strand | dichotomous | simple-branched | extant | 10 | ||
| Glossopteris | Glossopteridales | smooth | frequent | 2 | present | dichotomous | absent | Permian | 10 | |
| Gangamopteris | Glossopteridales | smooth | frequent | 1 | multistrand | dichotomous | absent | Permian | 10 | |
| Gigantonoclea | Gigantopteridales | frequent | 4 | multi-strand | unbranched | absent, branched | Permian | 10 | ||
| Delnortea | Gigantopteridales | frequent | 4 | multi-strand | unbranched | absent | Permian | 10 | ||
| Sagenopteris | Caytoniales | frequent | 2 | present | dichotomous | absent | Jurassic | 10 | ||
| Sanmiguelia | Uncertain | variable | 4 | absent | variable | absent | Triassic | 10 | ||
| Marcouia | Uncertain | frequent | 2 | present | dichotomous | absent | Triassic | 10 | ||
| Furcula | Uncertain | frequent | 3–4 | present | excurrent/dichotomous | present | Triassic | 10 | ||
| Pannaulika | Uncertain | frequent | 4 | present | excurrent | present | Triassic | 10 | ||
| Myrtophyllum geinitzii | Angiosperms | smooth | frequent | present | 4 | present | dichotomous | Cretaceous | 11,12 | |
| Myrtophyllum angustum | Angiosperms | present | 11 | |||||||
| Grevilleophyllum constans | Angiosperms | present | 11 | |||||||
| Eucalyptophyllum oblongifolium | Angiosperms | present | 11 | |||||||
| Eucalyptolaurus depreii | Angiosperms | smooth | frequent | present | 3 | present | brochidodromous | absent | Cretaceous | 11 |
| Eucalyptolaurus | Angiosperms | smooth | frequent | present | 3 | present | brochidodromous | absent | Cretaceous | |
| Callianthus | Angiosperms | smooth | frequent | present | 1 | absent | dichotomous | absent | Cretaceous | |
| Our leaf | smooth | frequent | present | 2? | present | unbranched | rare | Jurassic |
Our survey of fossil and extant taxa with reticulate venation indicates that, besides the implication given by reticulate venation, the occurrence of intramarginal vein appears to be restricted to angiosperms (Table 1). Therefore the occurrence of intramarginal vein in our new leaf seems to underscore its possibility of an angiosperm. This inference is further strengthened by the occurrence of freely ending veinlet in areole, which, although not strictly restricted to angiosperms, is only seen in angiosperms, Gnetales, and fossil taxa of uncertain affinity. Taking all together, despite its Jurassic age and partial preservation, our new leaf with reticulate venation, intramarginal vein, and freely ending veinlet suggests that, unlike widely-believed, angiosperms are more likely to be a truthful existence in the Jurassic, in line with previous reports of Jurassic flowers [1-4, 13] and implication given by molecular clock studies [14].
Acknowledgement
This research was supported by the National Natural Science Foundation of China (42288201, 41688103, 91514302), Strategic Priority Research Program (B) of Chinese Academy of Sciences (XDB26000000), and Natural Science Foundation of Ningxia (2021AAC03471).
References
- Wang X (2010) Schmeissneria: An angiosperm from the Early Jurassic. Journal of Systematics and Evolution 48: 326-335. [crossref]
- Wang X, Duan S, Geng B, Cui J, Yang Y (2007) Schmeissneria: A missing link to angiosperms? BMC Evolutionary Biology 7. [crossref]
- Fu Q, Jose BD, Mike P, Manuel GÁ, Zhong JL et al. (2018) An unexpected noncarpellate epigynous flower from the Jurassic of China. eLife 7. [crossref]
- Fu Q, Diez JB, Pole M, García ÁM, Wang X (2020) Nanjinganthus is an angiosperm, isn’t it?. China Geology 3: 359-361.
- Tanner LH, Wang X, Morabito AC (2012) Fossil charcoal from the Middle Jurassic of the Ordos Basin, China and its paleoatmospheric implications. Geoscience Frontiers 3: 493-502.
- Wang X (1995) Study on the middle Jurassic flora of Tongchuan, Shaanxi Province. Chinese Journal of Botany 37: 81-88.
- Deng S, Fang L, Lu Y, Fan R, Yuan X (2010) The Mesozoic stratigraphy of the Ordos Basin. (Shehong, Sichuan, China).
- Deng S, et al. (2003) Stratum introduction. Petroleum Industry Press, Beijing.
- Golovneva LB, Grabovskiy AA (2019) The genus Hausmannia (Dipteridaceae) in the Cretaceous of the North-East of Russia and its paleobiogeographic implications. Cretaceous Research 93: 22-32.
- Trivett ML, Pigg KB (1996) A survey of reticulate venation among fossil and living land plants, in ” Flowering plant origin, evolution and phylogeny ” (eds) Taylor DW, Hickey LJ, Ch. 2: 8-31 (Chapman and Hall).
- Coiffard C, Gomez B, Thiébaut M, Kvacek J, Thévenard F, et al. (2009) Intramarginal veined Lauracease leaves from the Albian-Cenomanian of Charente-Maritime (western France). Palaeontology 52: 323-336.
- Gall L (2022) Paleobotany Division, Yale Peabody Museum. Yale University Peabody Museum. Occurrence dataset.
- Han L, Zhao Y, Zhao M, Sun J, Sun B, Wang, X (2023) New fossil evidence suggests that angiosperms flourished in the Middle Jurassic. Life 13.
- Li H, et al. (2019) Origin of angiosperms and the puzzle of the Jurassic gap. Nature Plants 5: 461-470.